Blavatnik Institute, Department of Genetics, Harvard Medical School, Boston, United States.
Elife. 2022 May 16;11:e78944. doi: 10.7554/eLife.78944.
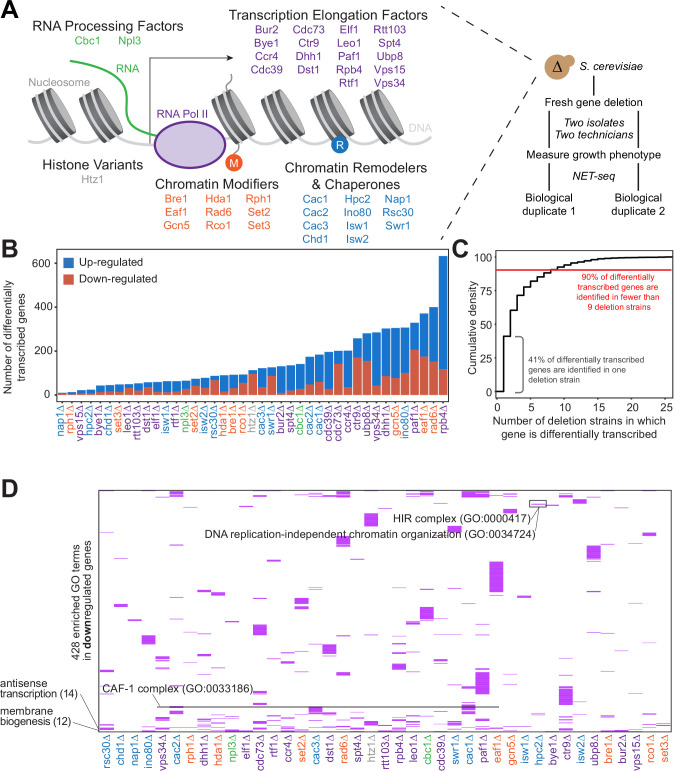
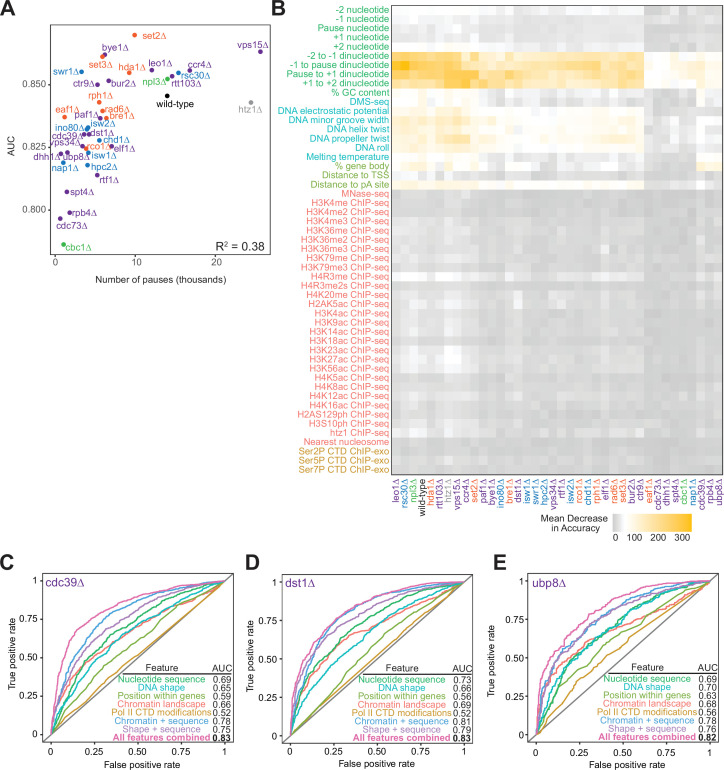
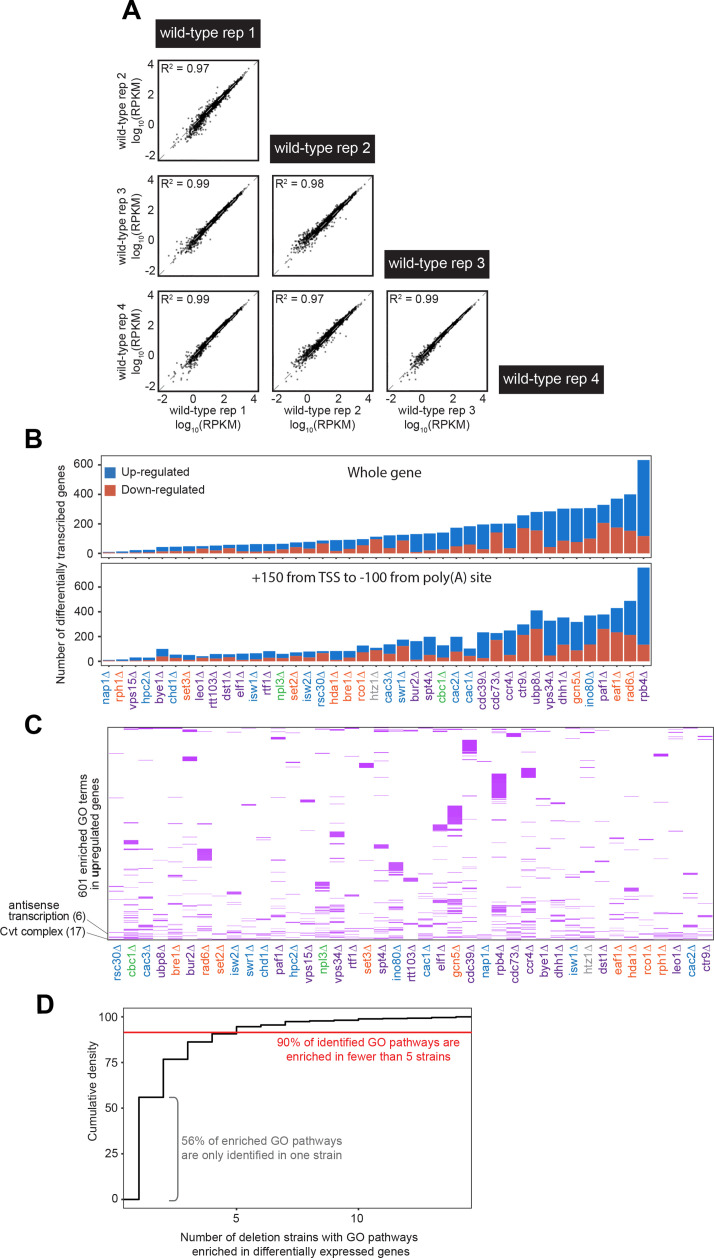
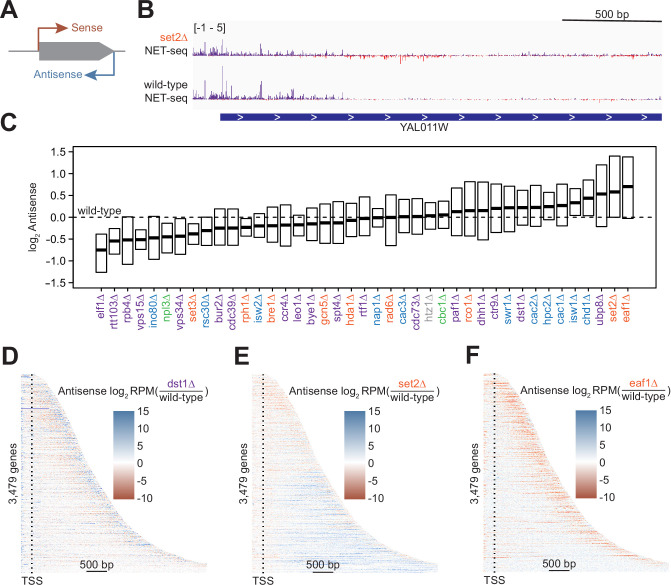
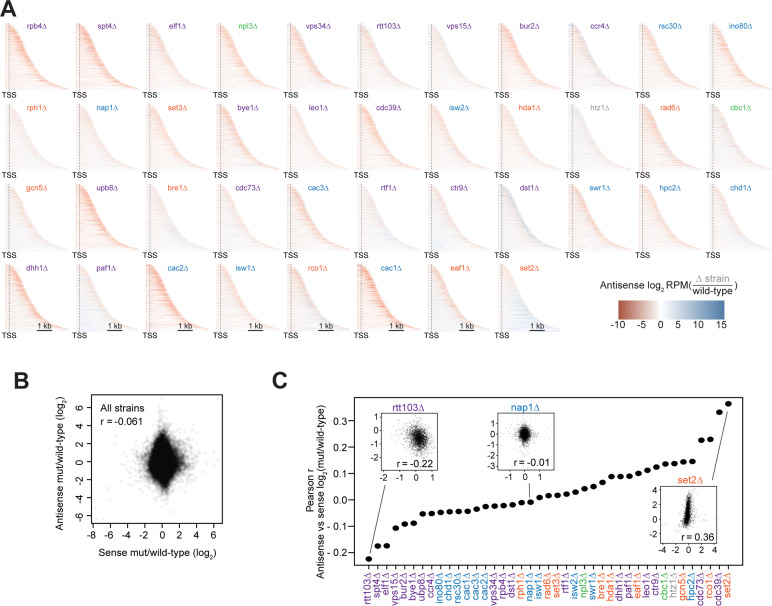
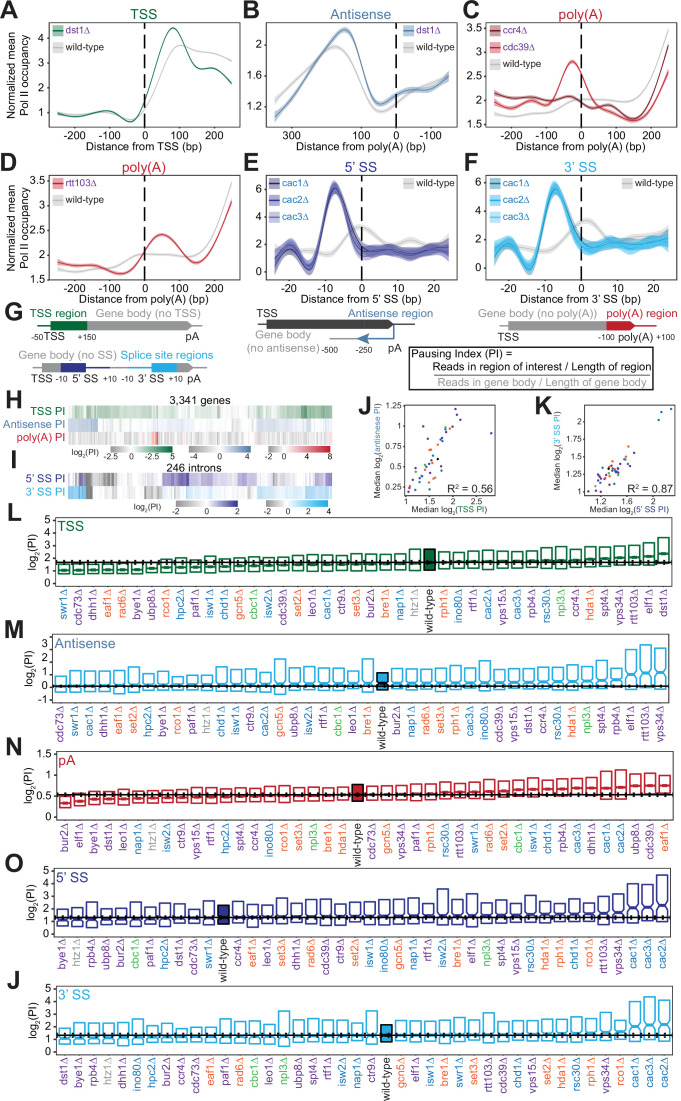
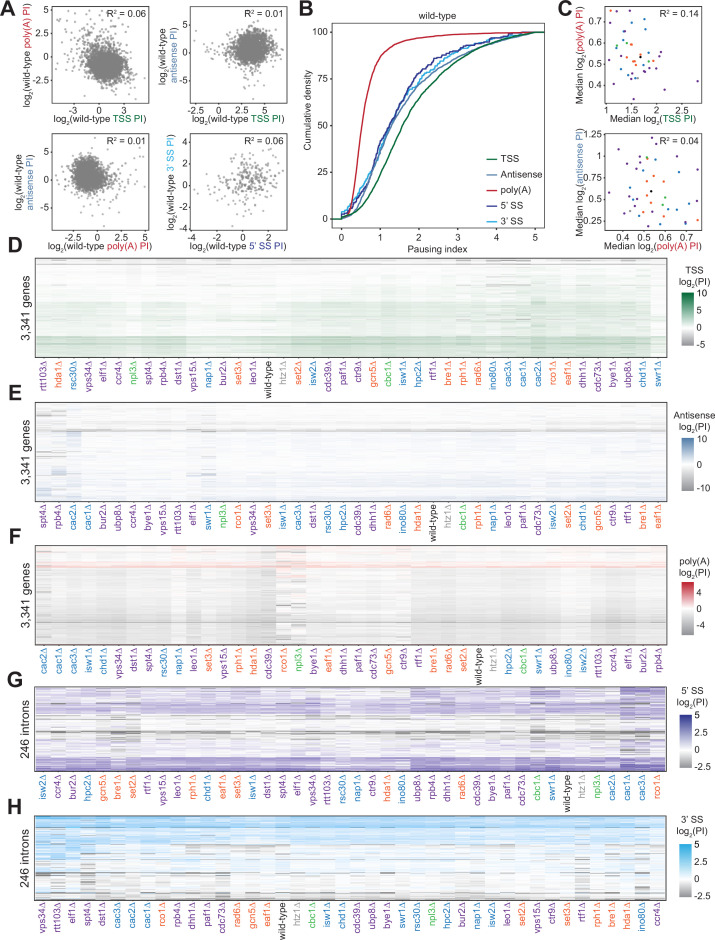
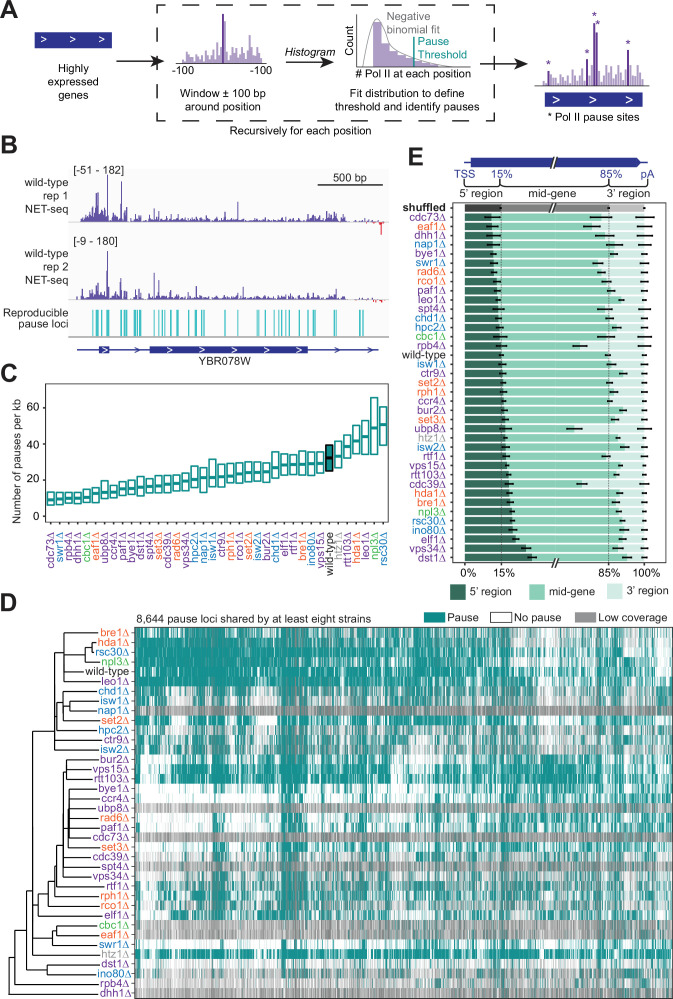
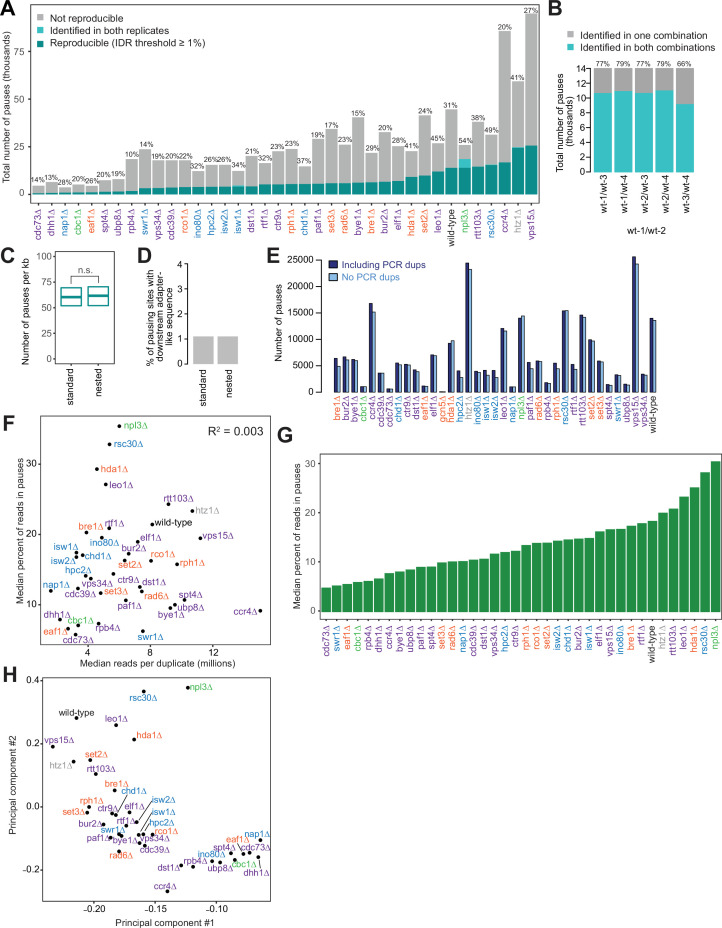
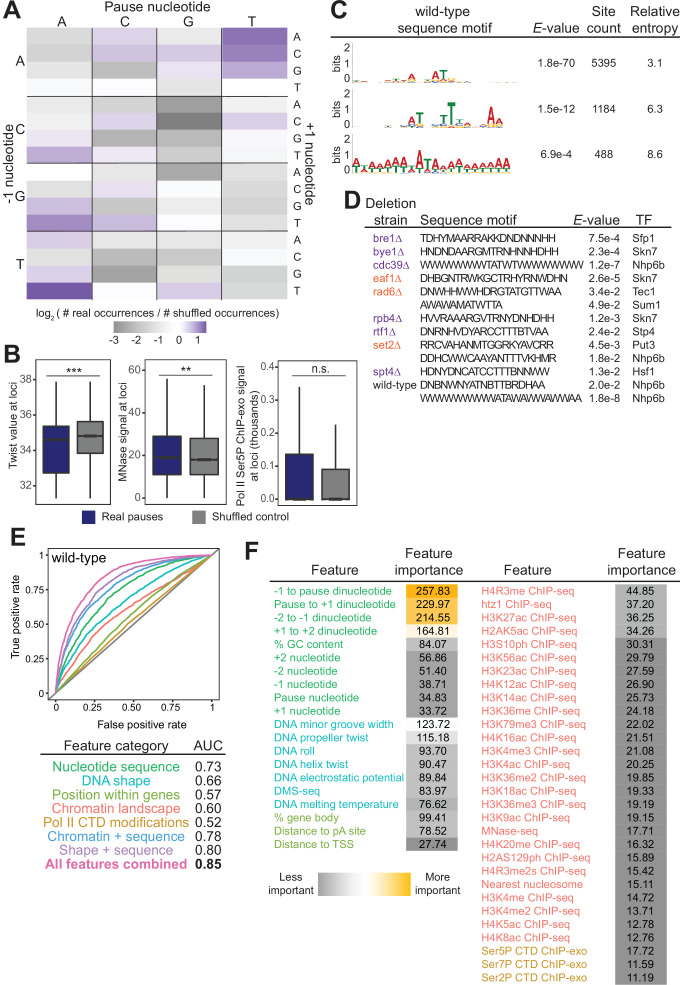
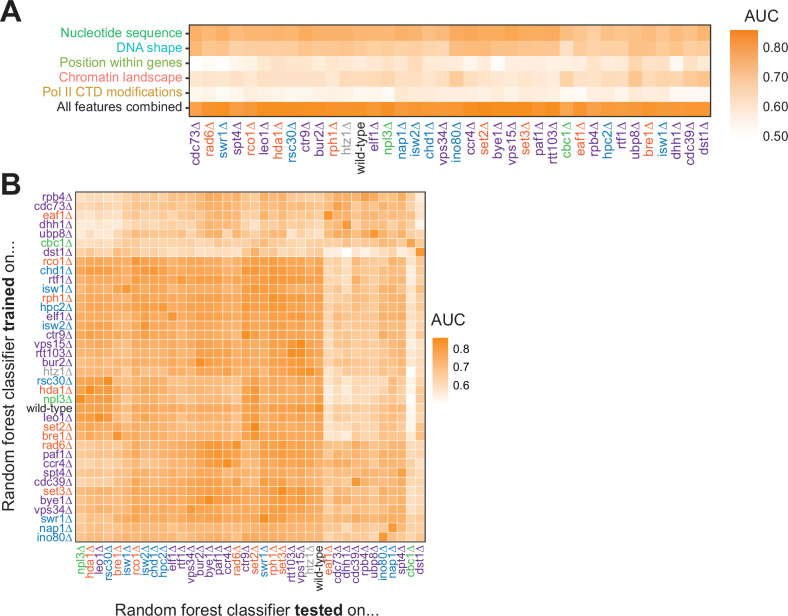
Understanding the complex network that regulates transcription elongation requires the quantitative analysis of RNA polymerase II (Pol II) activity in a wide variety of regulatory environments. We performed native elongating transcript sequencing (NET-seq) in 41 strains of lacking known elongation regulators, including RNA processing factors, transcription elongation factors, chromatin modifiers, and remodelers. We found that the opposing effects of these factors balance transcription elongation and antisense transcription. Different sets of factors tightly regulate Pol II progression across gene bodies so that Pol II density peaks at key points of RNA processing. These regulators control where Pol II pauses with each obscuring large numbers of potential pause sites that are primarily determined by DNA sequence and shape. Antisense transcription varies highly across the regulatory landscapes analyzed, but antisense transcription in itself does not affect sense transcription at the same locus. Our findings collectively show that a diverse array of factors regulate transcription elongation by precisely balancing Pol II activity.
理解调控转录延伸的复杂网络需要在广泛的调控环境中对 RNA 聚合酶 II(Pol II)的活性进行定量分析。我们在 41 株缺乏已知延伸调控因子的菌株中进行了天然延伸转录测序(NET-seq),这些因子包括 RNA 加工因子、转录延伸因子、染色质修饰因子和重塑因子。我们发现,这些因子的相反作用平衡了转录延伸和反义转录。不同的因子集严格调控 Pol II 在基因体中的进展,使得 Pol II 在 RNA 加工的关键点处达到峰值。这些调节剂通过每个点的暂停来控制 Pol II 的位置,从而掩盖了大量主要由 DNA 序列和形状决定的潜在暂停点。在分析的调控景观中,反义转录变化很大,但反义转录本身不会影响同一基因座上的有意义转录。我们的研究结果表明,各种因素通过精确平衡 Pol II 活性来调节转录延伸。