Department of Orthopaedic Surgery, University of California, San Francisco, San Francisco, United States.
Elife. 2022 Jun 6;11:e66005. doi: 10.7554/eLife.66005.
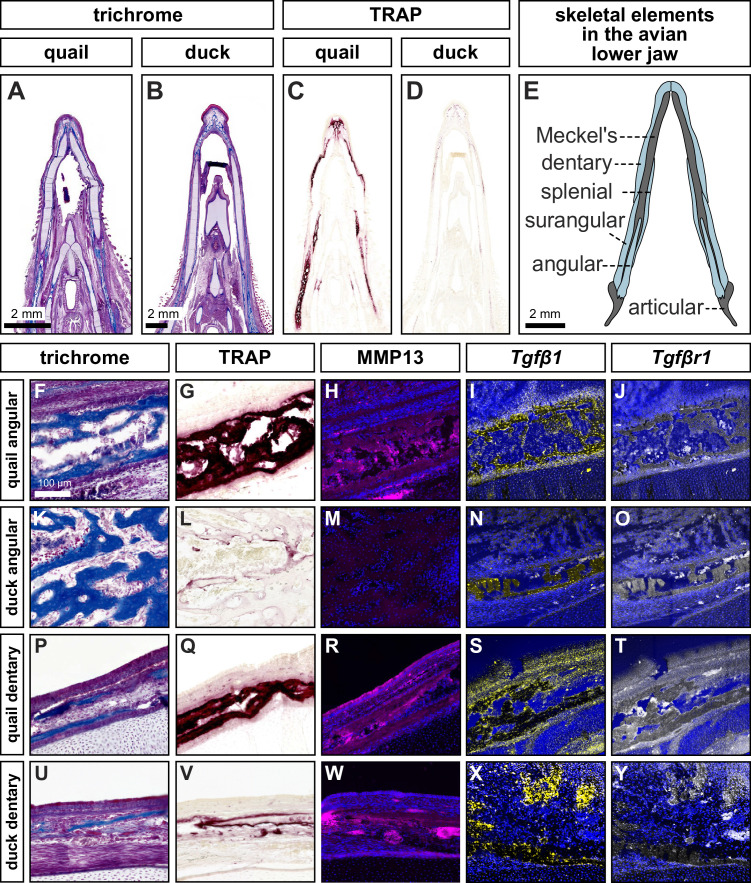
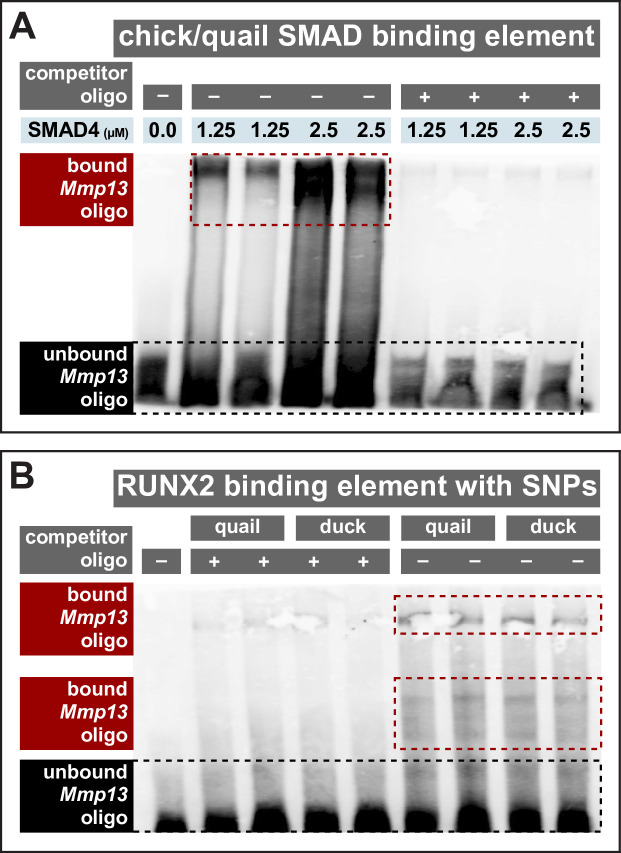
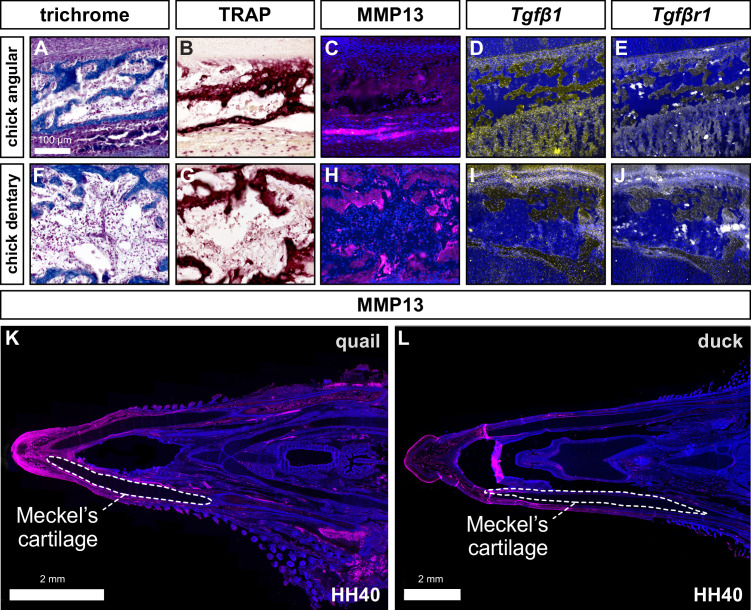
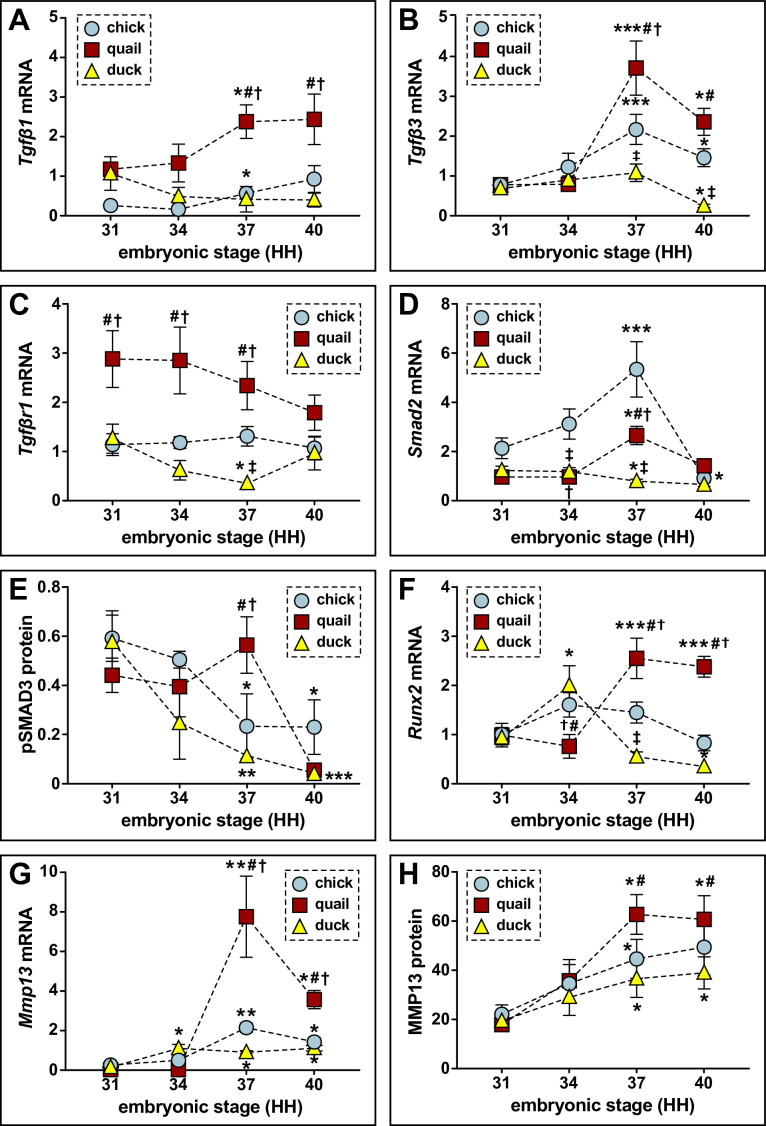
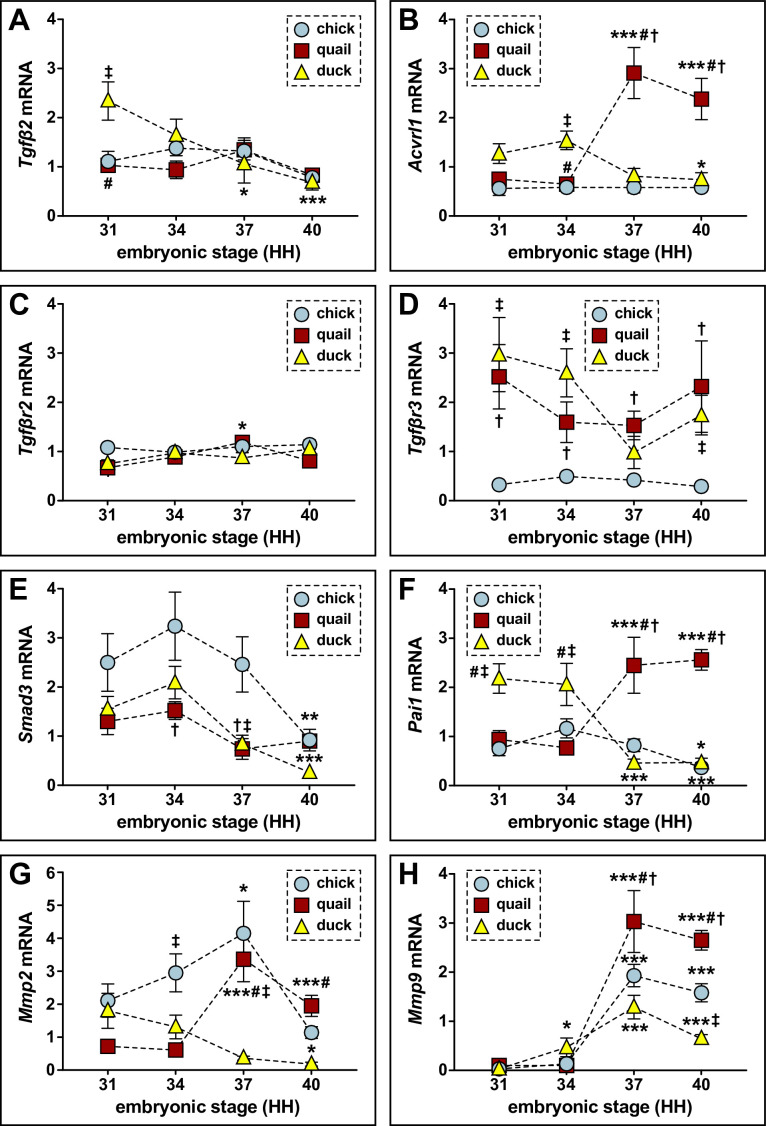
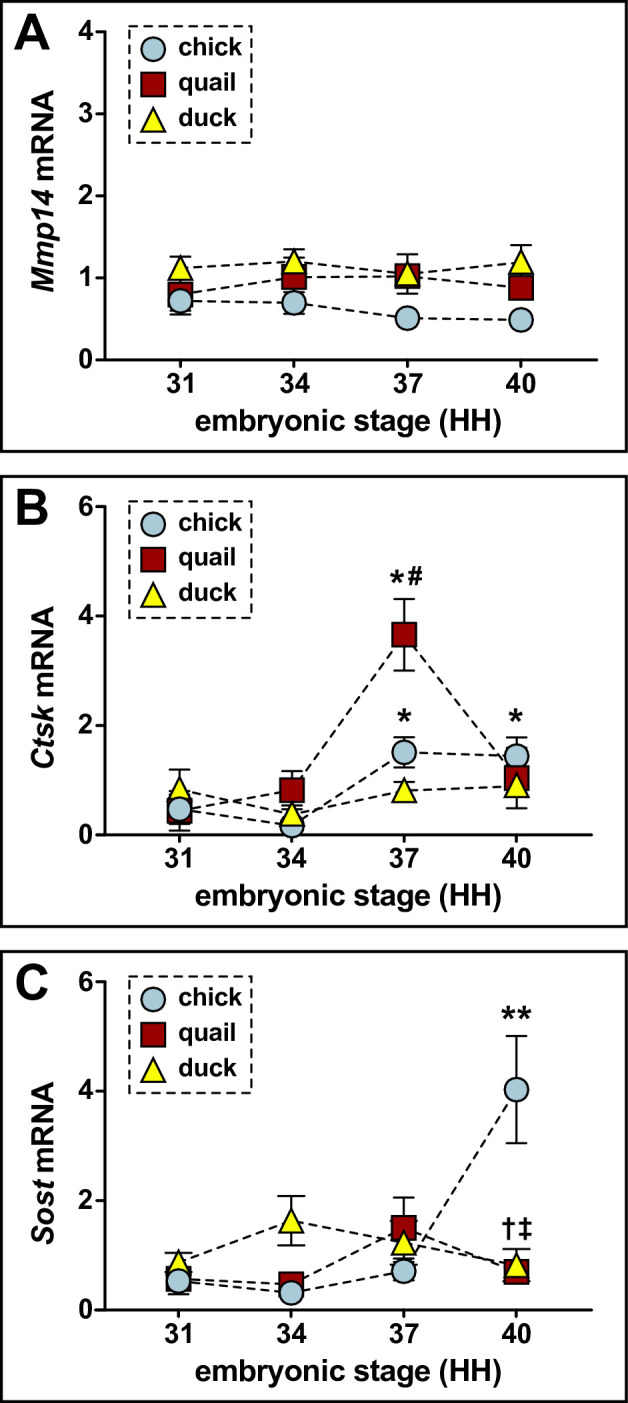
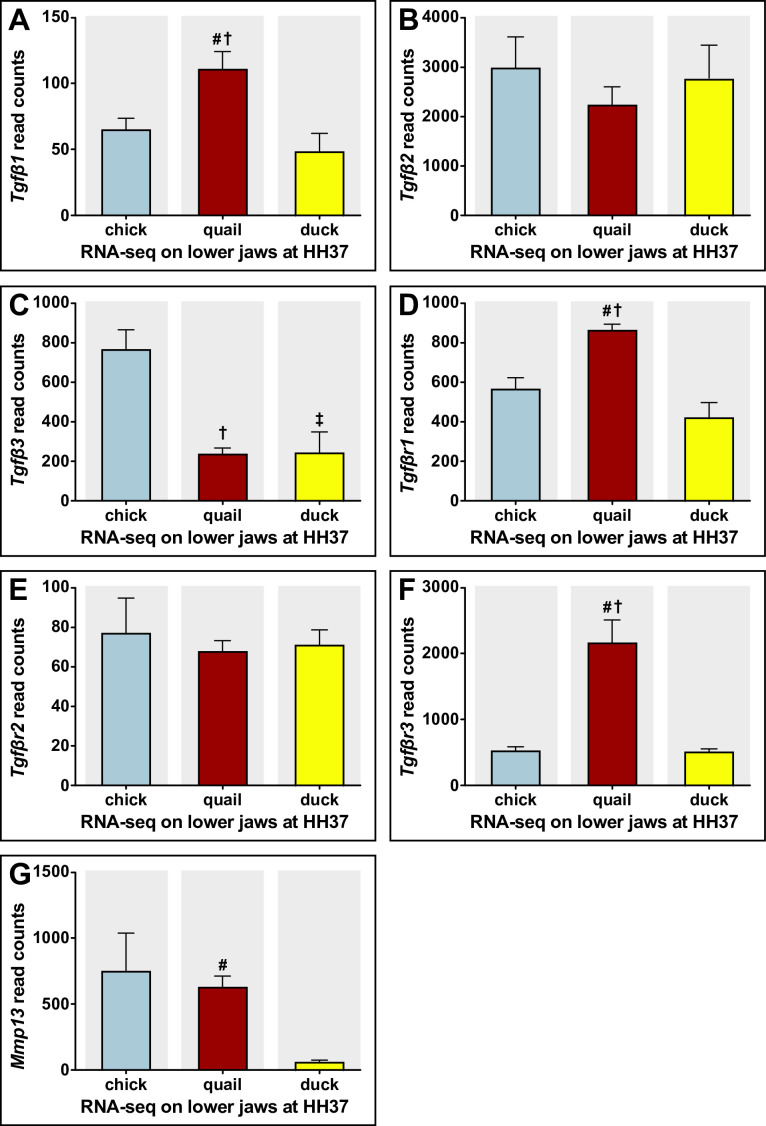
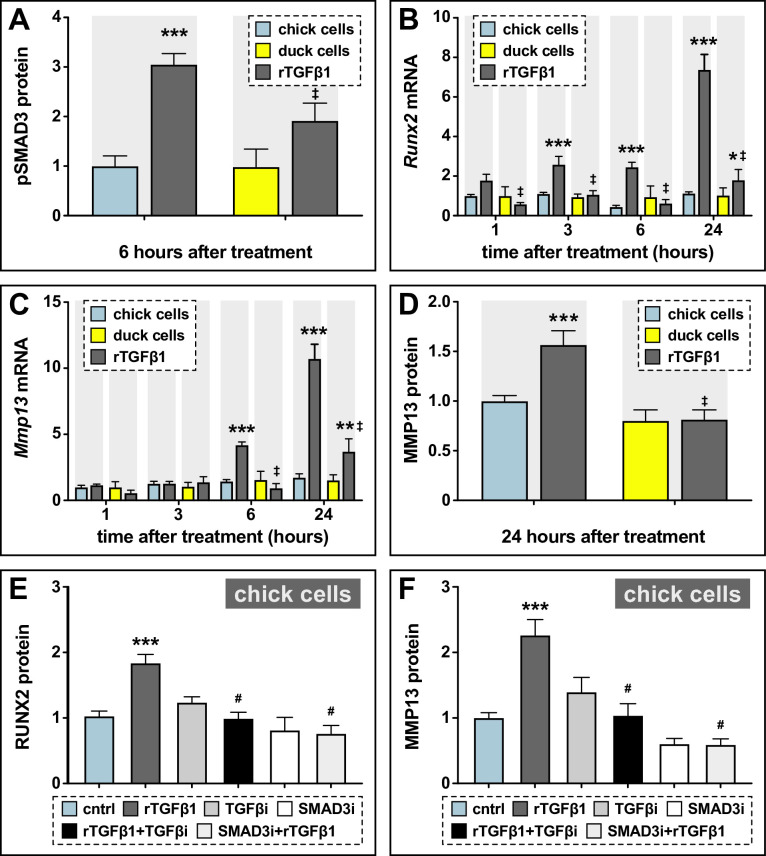
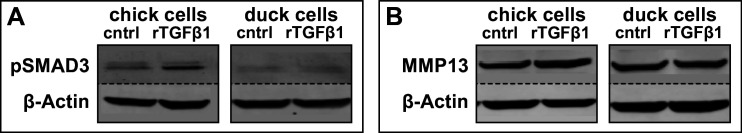
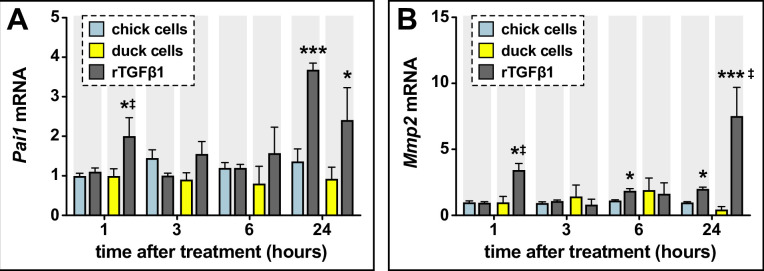
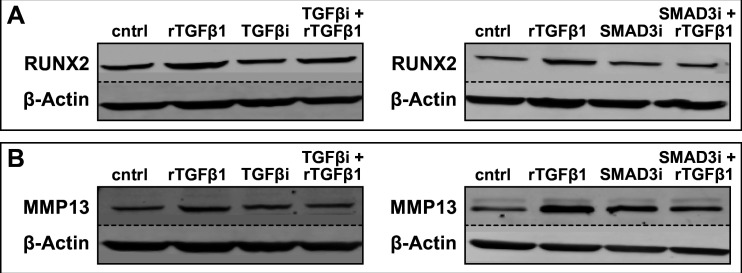
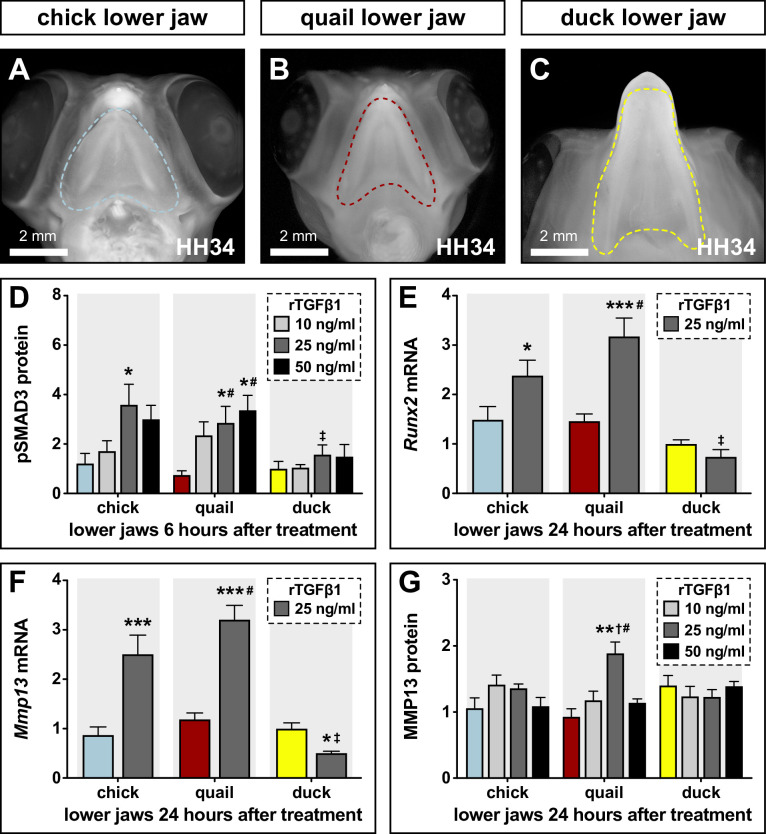
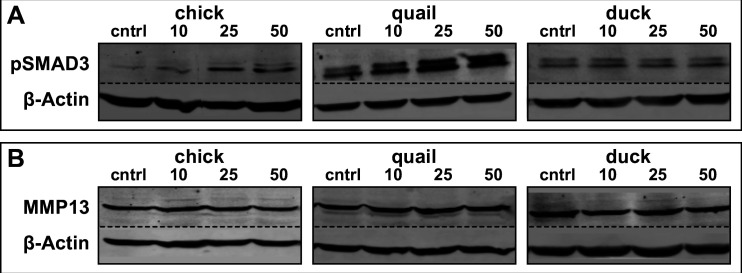
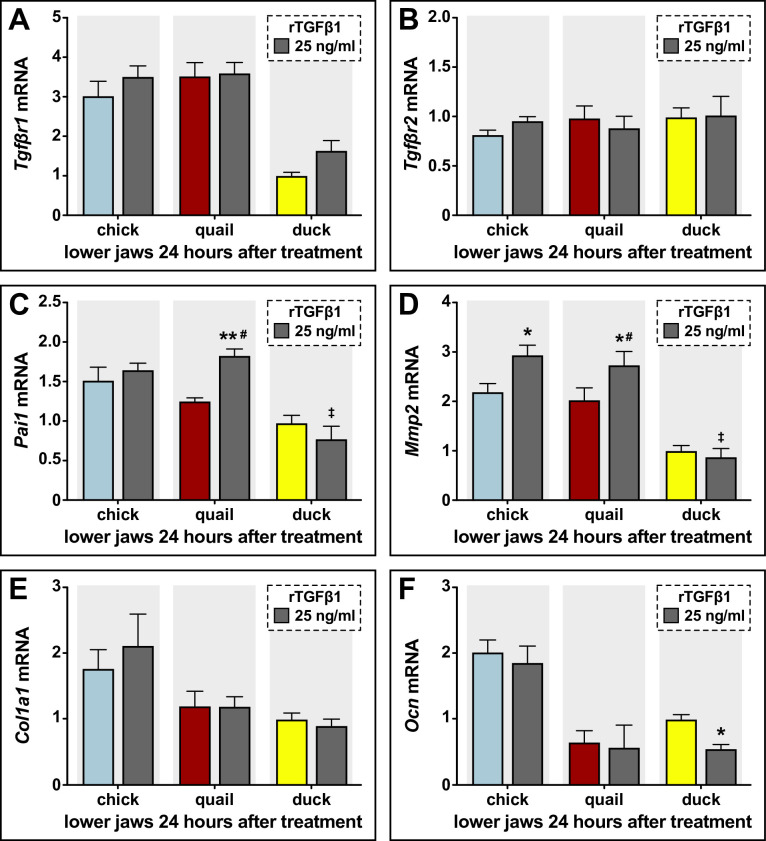
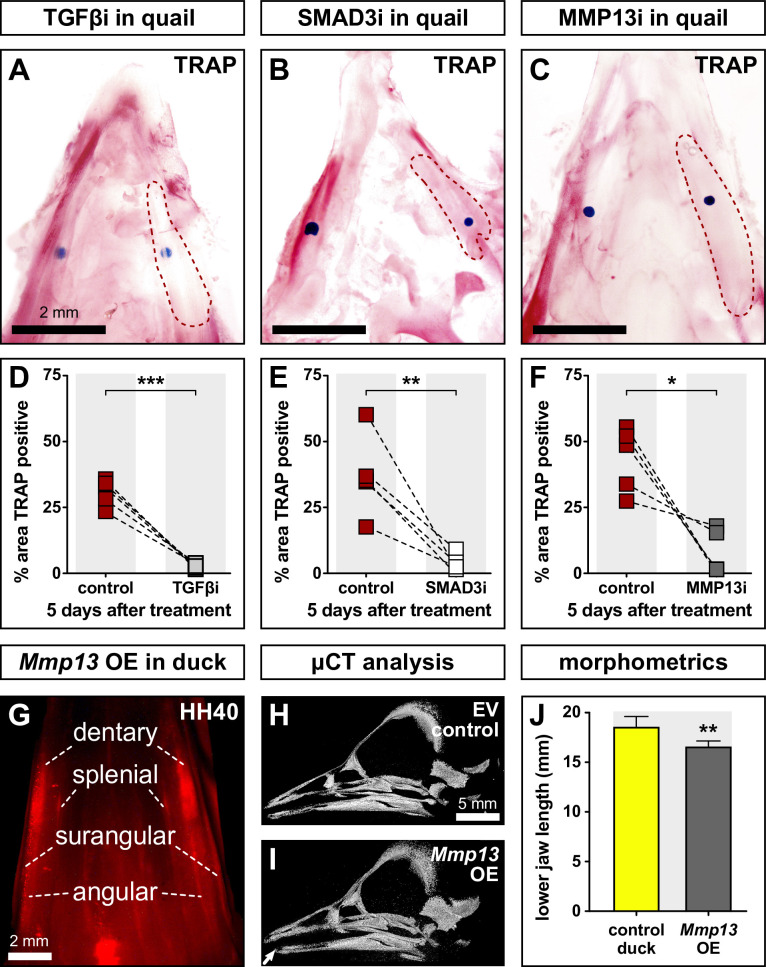
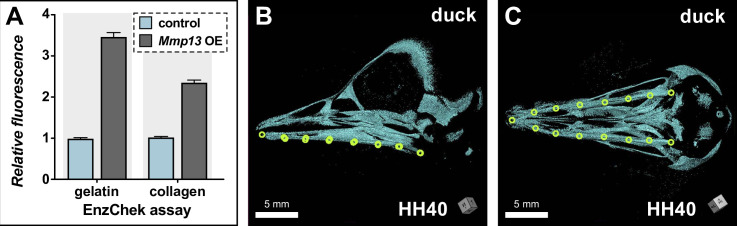
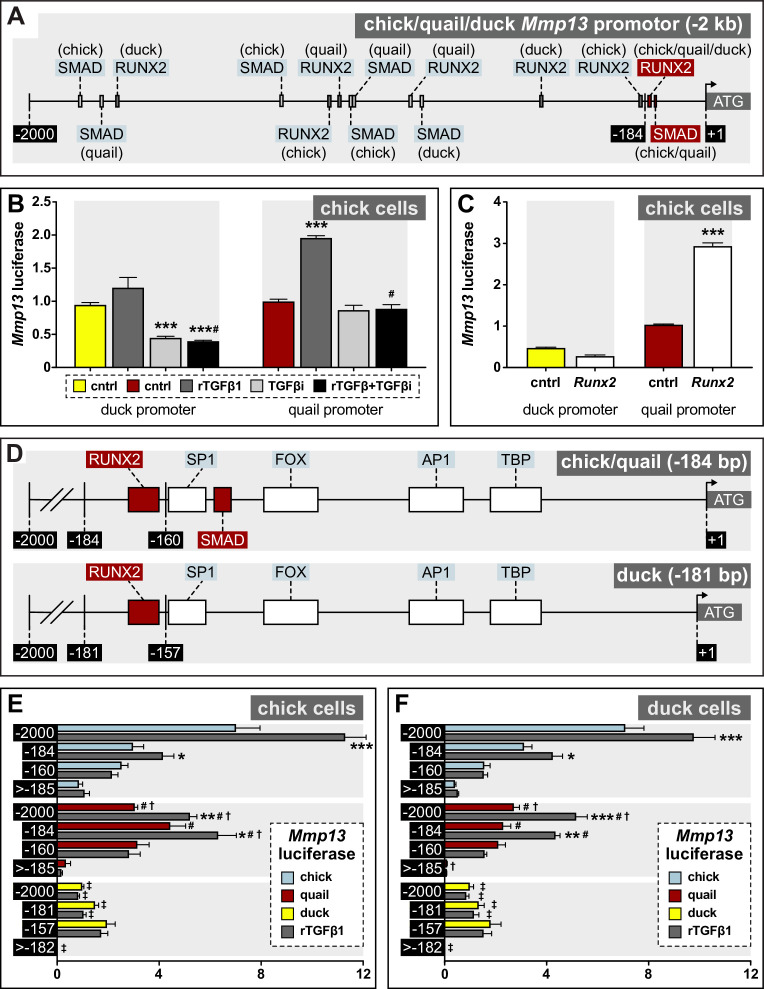
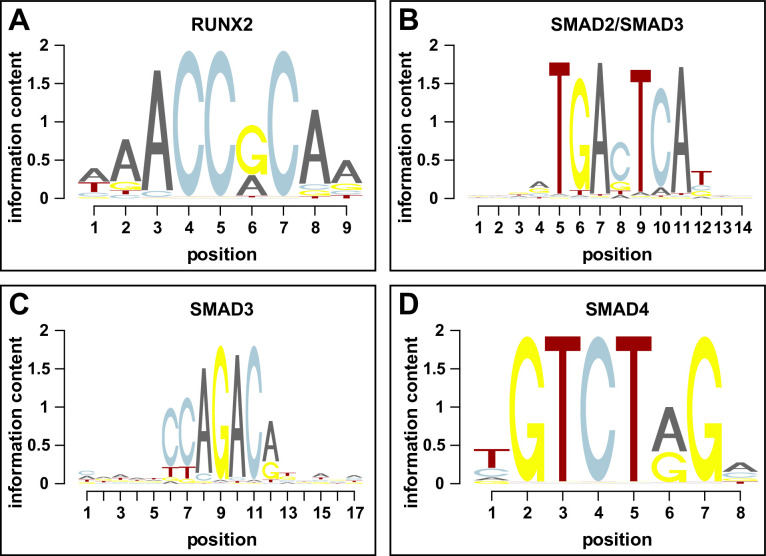
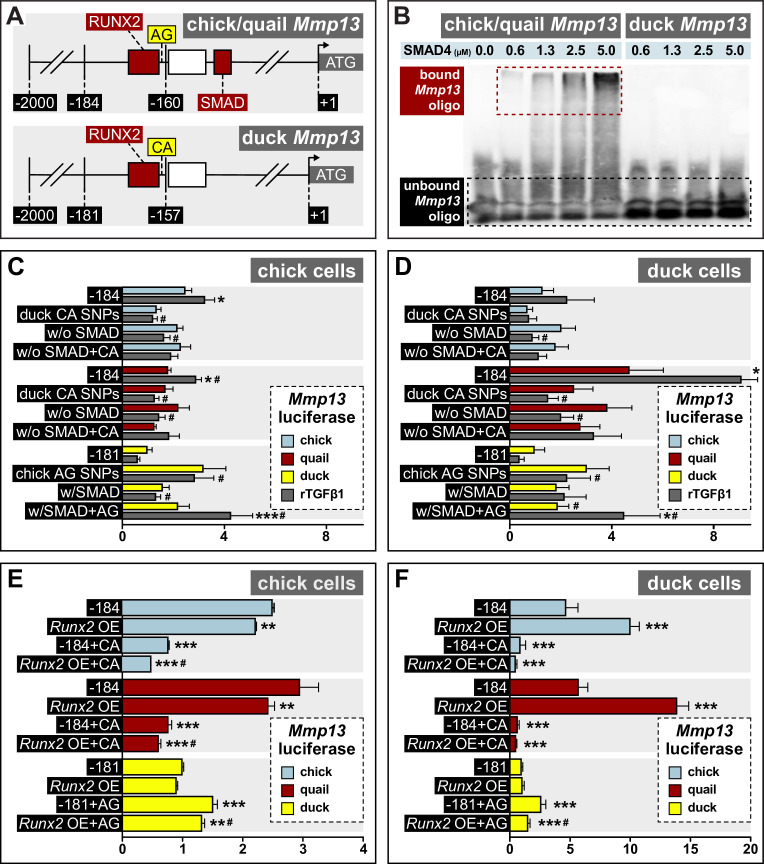
Precise developmental control of jaw length is critical for survival, but underlying molecular mechanisms remain poorly understood. The jaw skeleton arises from neural crest mesenchyme (NCM), and we previously demonstrated that these progenitor cells express more bone-resorbing enzymes including () when they generate shorter jaws in quail embryos versus longer jaws in duck. Moreover, if we inhibit bone resorption or we can increase jaw length. In the current study, we uncover mechanisms establishing species-specific levels of and bone resorption. Quail show greater activation of and sensitivity to transforming growth factor beta (TGFβ) signaling than duck; where intracellular mediators like SMADs and targets like () which bind , become elevated. Inhibiting TGFβ signaling decreases bone resorption, and overexpressing in NCM shortens the duck lower jaw. To elucidate the basis for this differential regulation, we examine the promoter. We discover a SMAD-binding element and single nucleotide polymorphisms (SNPs) near a RUNX2-binding element that distinguish quail from duck. Altering the SMAD site and switching the SNPs abolish TGFβ sensitivity in the quail promoter but make the duck promoter responsive. Thus, differential regulation of TGFβ signaling and promoter structure underlie avian jaw development and evolution.
精确的颌骨长度发育控制对生存至关重要,但相关的分子机制仍知之甚少。颌骨骨架来源于神经嵴间质(NCM),我们之前的研究表明,这些祖细胞在鹌鹑胚胎中生成较短的颌骨时,比在鸭胚胎中生成较长的颌骨时,表达更多的骨吸收酶,包括 ()。此外,如果我们抑制骨吸收或 (),可以增加颌骨长度。在本研究中,我们揭示了建立物种特异性 () 和骨吸收水平的机制。鹌鹑比鸭表现出更高的转化生长因子β(TGFβ)信号激活和敏感性;细胞内介质如 SMADs 和结合 () 的靶基因,如 (),会升高。抑制 TGFβ 信号可减少骨吸收,过表达 NCM 中的 ()可缩短鸭的下颌骨。为了阐明这种差异调控的基础,我们研究了 () 启动子。我们发现了一个 SMAD 结合元件和一个 RUNX2 结合元件附近的单核苷酸多态性(SNP),这将鹌鹑与鸭区分开来。改变 SMAD 位点并切换 SNP 会使鹌鹑 () 启动子失去 TGFβ 敏感性,但使鸭启动子具有反应性。因此,TGFβ 信号转导和 () 启动子结构的差异调控是鸟类颌骨发育和进化的基础。