MOE Laboratory of Biosystems Homeostasis & Protection of College of Life Sciences, Key Laboratory of Cardiovascular Intervention and Regenerative Medicine of Zhejiang Province of Sir Run Shaw Hospital, Zhejiang University, Hangzhou, Zhejiang 310058, China.
Cardiovascular Research Center, Interdepartmental Program in Vascular Biology and Therapeutics, Department of Internal Medicine, Yale University School of Medicine, New Haven, CT 06520.
Proc Natl Acad Sci U S A. 2022 Jun 28;119(26):e2202631119. doi: 10.1073/pnas.2202631119. Epub 2022 Jun 21.
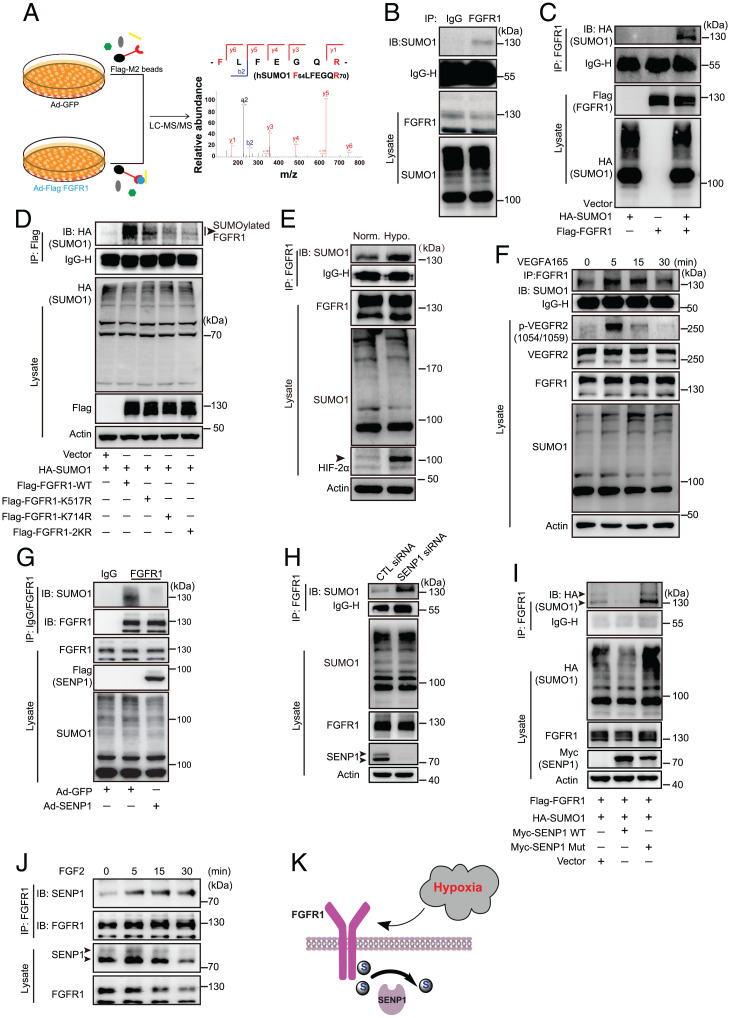
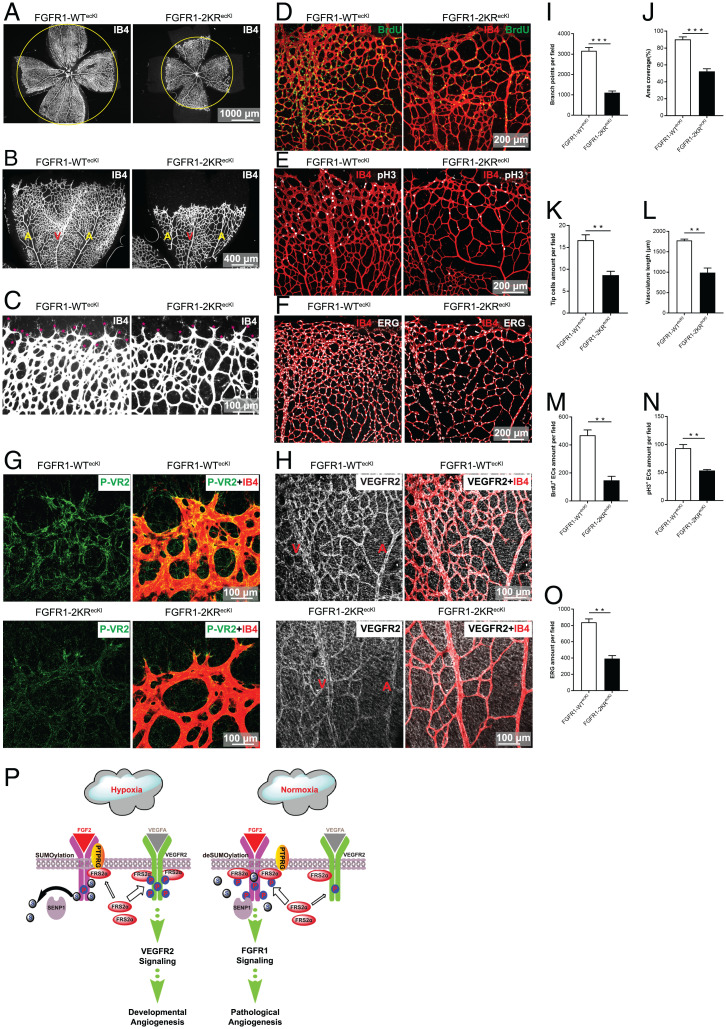
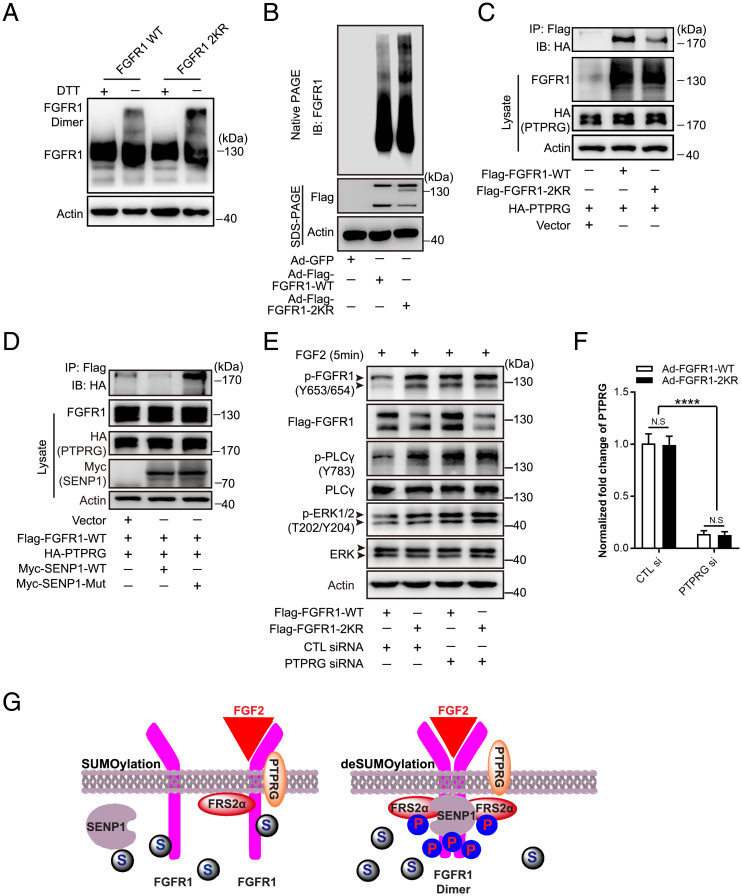
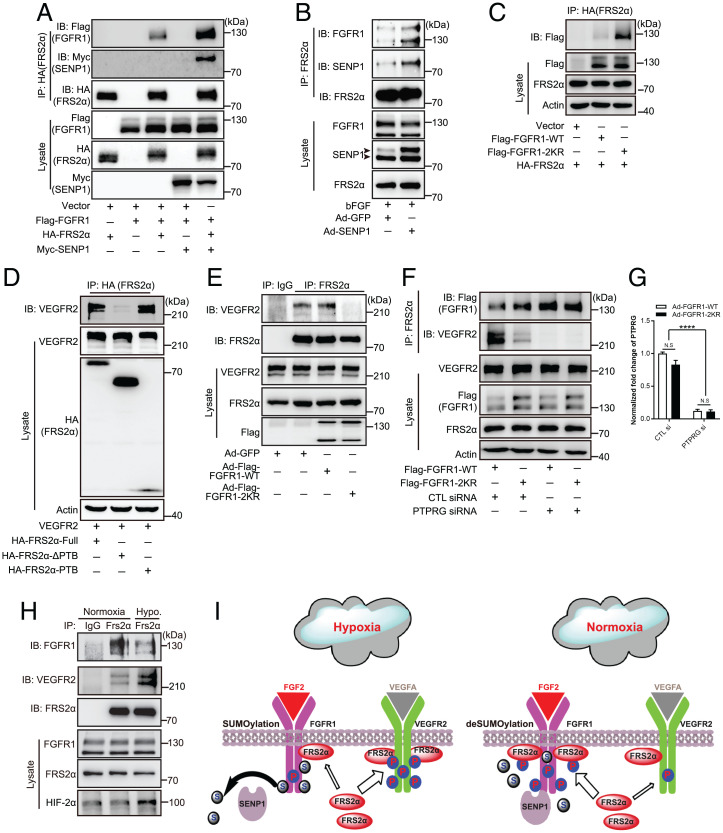
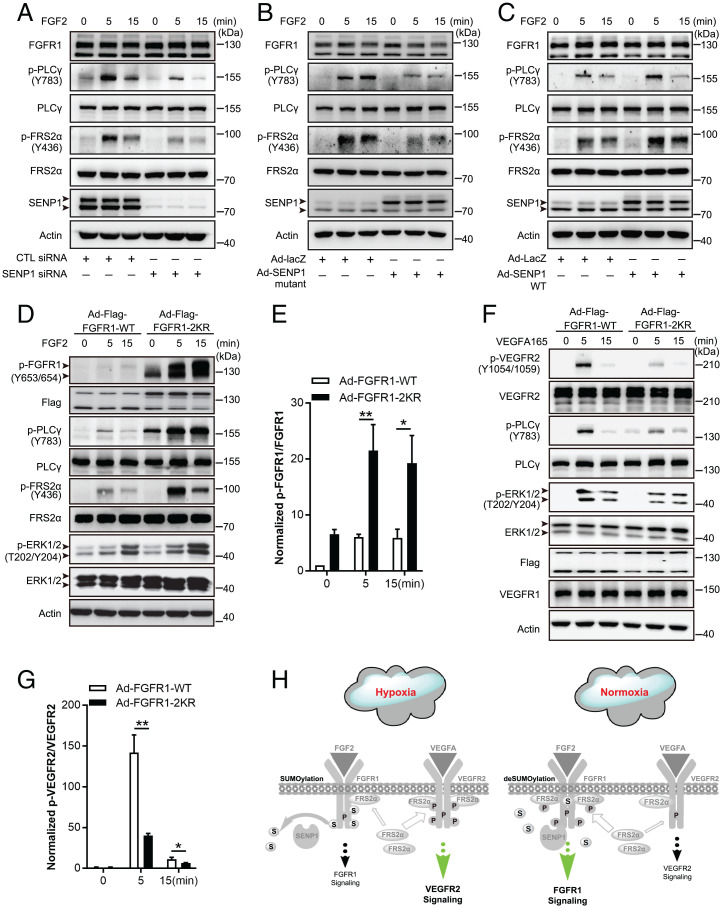
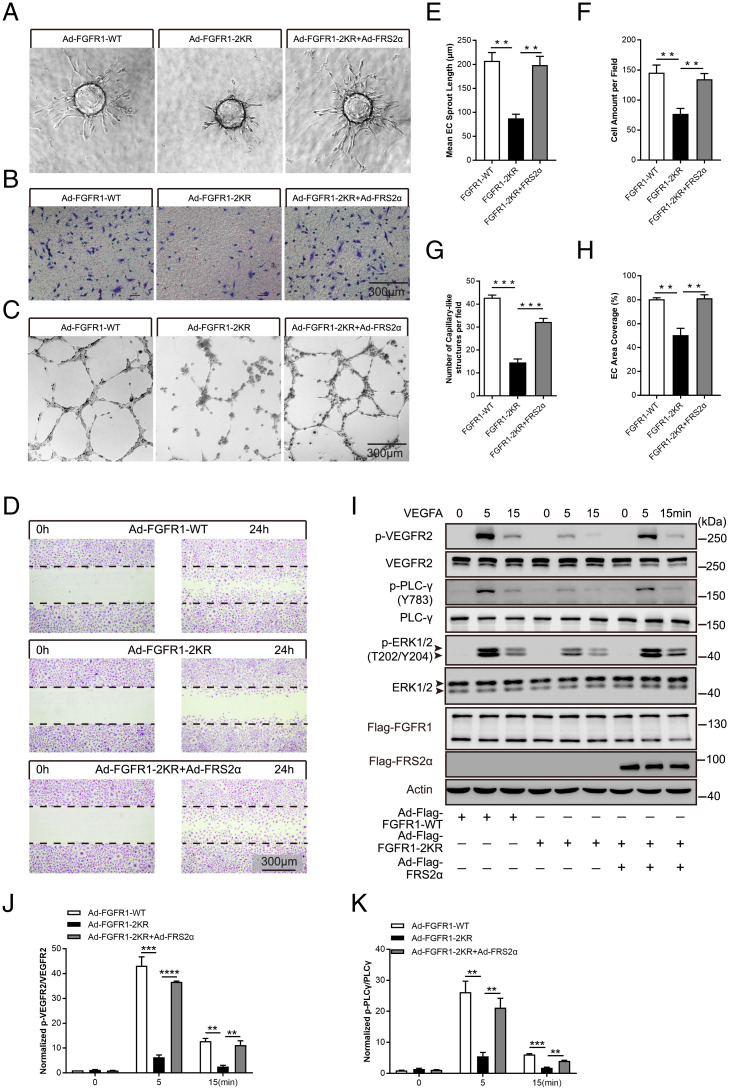
Angiogenesis contributes fundamentally to embryonic development, tissue homeostasis, and wound healing. Basic fibroblast growth factor (FGF2) is recognized as the first proangiogenic molecule discovered, and it facilitates angiogenesis by activating FGF receptor 1 (FGFR1) signaling in endothelial cells. However, the precise roles of FGFR and the FGF/FGFR signaling axis in angiogenesis remain unclear, especially because of the contradictory phenotypes of in vivo FGF and FGFR gene deficiency models. Our previous study results suggested a potential role of posttranslational small ubiquitin-like modifier modification (SUMOylation), with highly dynamic regulatory features, in vascular development and disorder. Here, we identified SENP1-regulated endothelial FGFR1 SUMOylation at conserved lysines responding to proangiogenic stimuli, while SENP1 functioned as the deSUMOylase. Hypoxia-enhanced FGFR1 SUMOylation restricted the tyrosine kinase activation of FGFR1 by modulating the dimerization of FGFR1 and FGFR1 binding with its phosphatase PTPRG. Consequently, it facilitated the recruitment of FRS2α to VEGFR2 but limited additional recruitment of FRS2α to FGFR1, supporting the activation of VEGFA/VEGFR2 signaling in endothelial cells. Furthermore, SUMOylation-defective mutation of FGFR1 resulted in exaggerated FGF2/FGFR1 signaling but suppressed VEGFA/VEGFR2 signaling and the angiogenic capabilities of endothelial cells, which were rescued by FRS2α overexpression. Reduced angiogenesis and endothelial sprouting in mice bearing an endothelial-specific, FGFR1 SUMOylation-defective mutant confirmed the functional significance of endothelial FGFR1 SUMOylation in vivo. Our findings identify the reversible SUMOylation of FGFR1 as an intrinsic fine-tuned mechanism in coordinating endothelial angiogenic signaling during neovascularization; SENP1-regulated FGFR1 SUMOylation and deSUMOylation controls the competitive recruitment of FRS2α by FGFR1 and VEGFR2 to switch receptor-complex formation responding to hypoxia and normoxia angiogenic environments.
血管生成对于胚胎发育、组织稳态和伤口愈合至关重要。碱性成纤维细胞生长因子(FGF2)被认为是第一个被发现的促血管生成分子,它通过在血管内皮细胞中激活 FGF 受体 1(FGFR1)信号来促进血管生成。然而,FGFR 和 FGF/FGFR 信号轴在血管生成中的确切作用仍不清楚,尤其是由于体内 FGF 和 FGFR 基因缺失模型的表型相互矛盾。我们之前的研究结果表明,翻译后小泛素样修饰物修饰(SUMOylation)具有高度动态的调节特征,可能在血管发育和紊乱中发挥作用。在这里,我们鉴定了 SENP1 调节的内皮 FGFR1 SUMOylation 在内皮细胞中对促血管生成刺激的反应,而 SENP1 作为去 SUMO 酶发挥作用。缺氧增强了 FGFR1 SUMOylation,通过调节 FGFR1 二聚化和 FGFR1 与磷酸酶 PTPRG 的结合,限制了 FGFR1 的酪氨酸激酶激活。因此,它促进了 FRS2α 向 VEGFR2 的募集,但限制了 FRS2α 向 FGFR1 的额外募集,从而支持内皮细胞中 VEGFA/VEGFR2 信号的激活。此外,FGFR1 的 SUMOylation 缺陷突变导致 FGF2/FGFR1 信号过度激活,但抑制了 VEGFA/VEGFR2 信号和内皮细胞的血管生成能力,而过表达 FRS2α 可以挽救这种情况。在携带内皮细胞特异性 FGFR1 SUMOylation 缺陷突变的小鼠中,血管生成减少和内皮细胞出芽证实了内皮细胞 FGFR1 SUMOylation 在体内的功能意义。我们的研究结果确定了 FGFR1 的可逆 SUMOylation 是协调血管生成信号的内在精细调节机制,在血管新生过程中,SENP1 调节的 FGFR1 SUMOylation 和去 SUMOylation 控制了 FRS2α 通过 FGFR1 和 VEGFR2 的竞争募集,以响应缺氧和常氧血管生成环境切换受体复合物的形成。