The Eli and Edythe Broad Center of Regeneration Medicine and Stem Cell, University of California, San Francisco, San Francisco, United States.
Department of Urology, University of California, San Francisco, San Francisco, United States.
Elife. 2022 Jul 7;11:e78829. doi: 10.7554/eLife.78829.
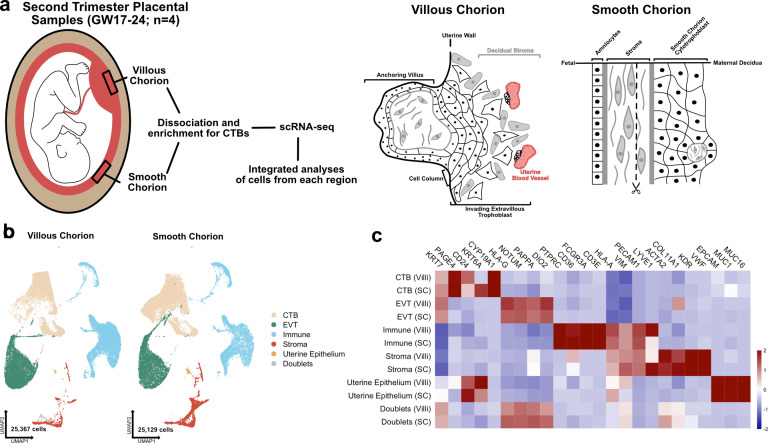
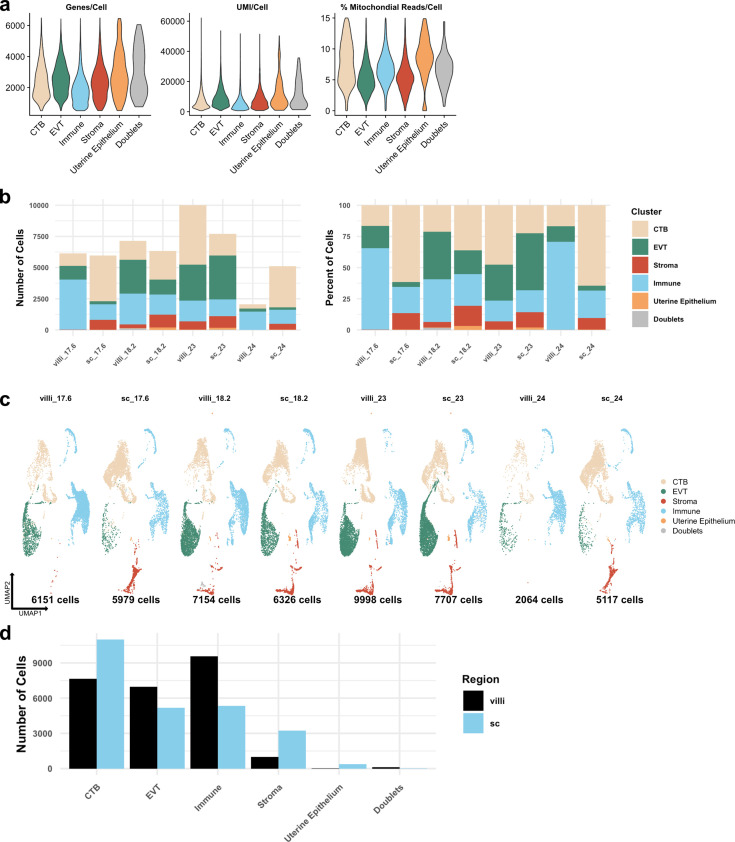
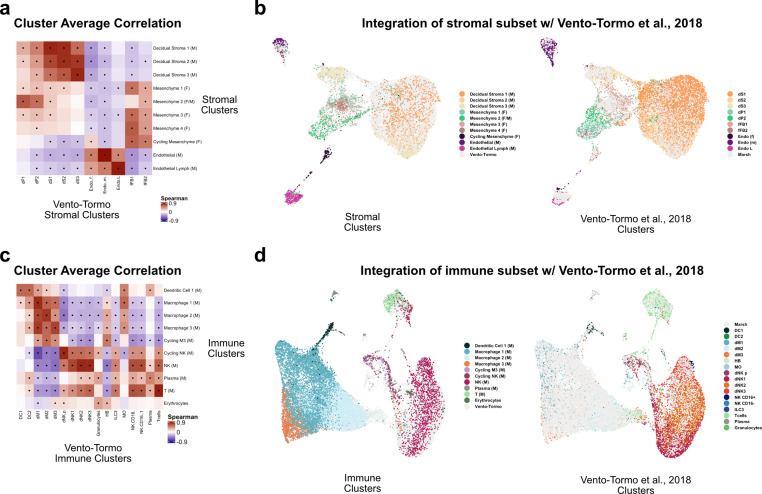
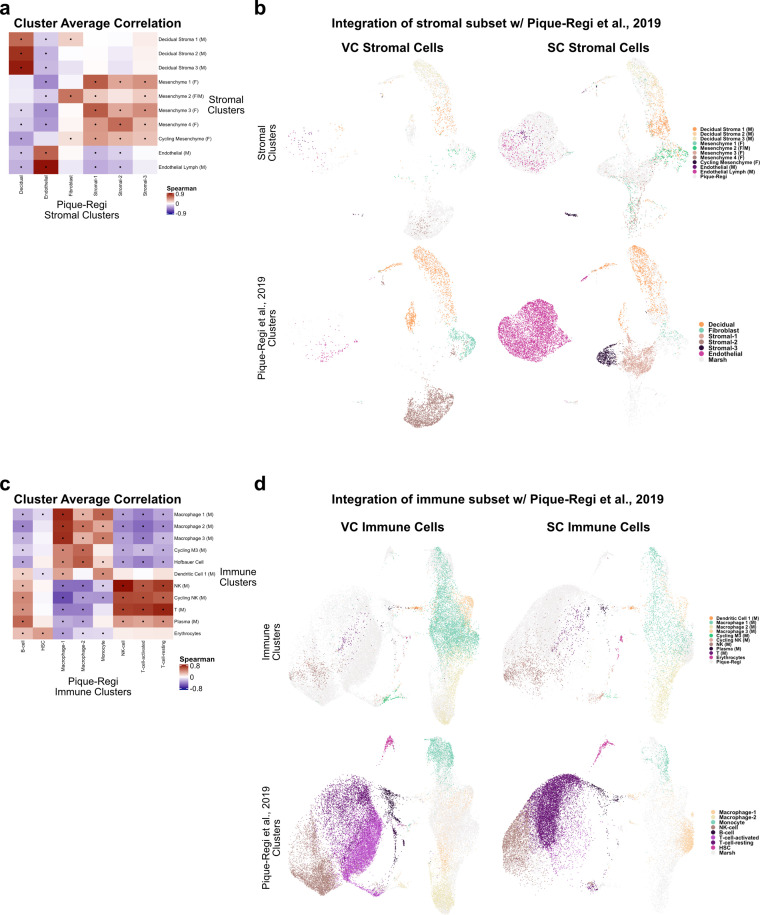
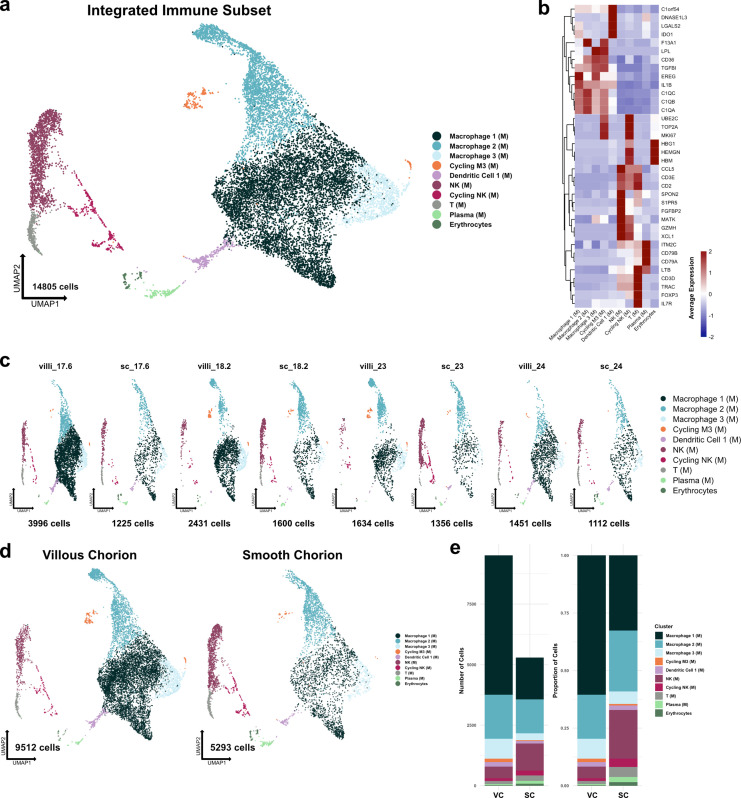
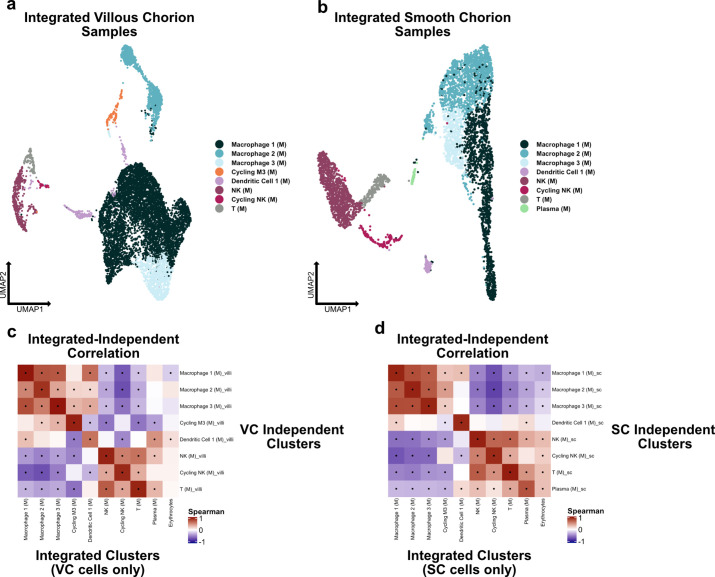
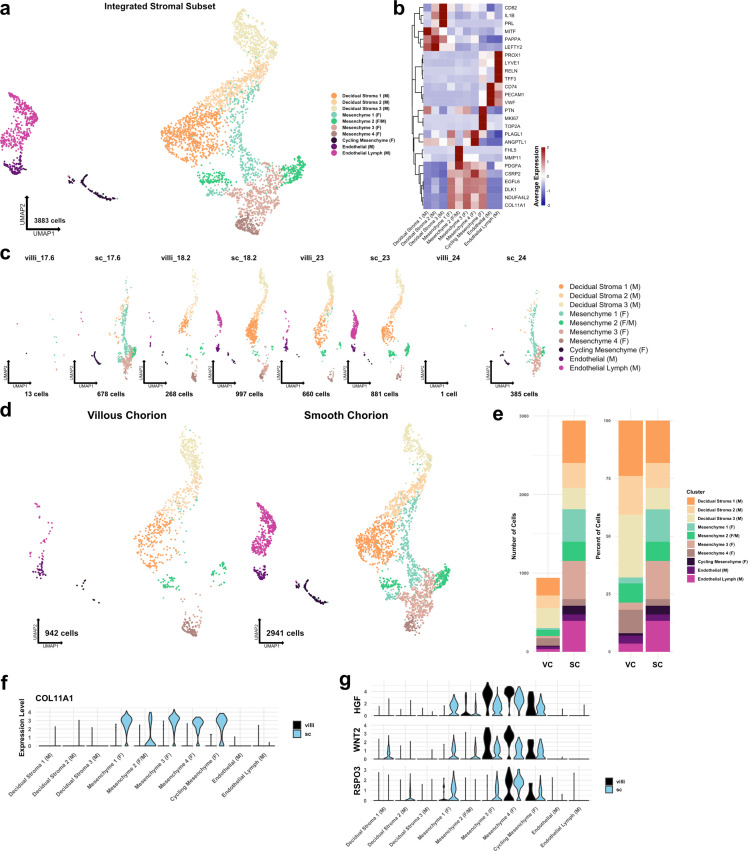
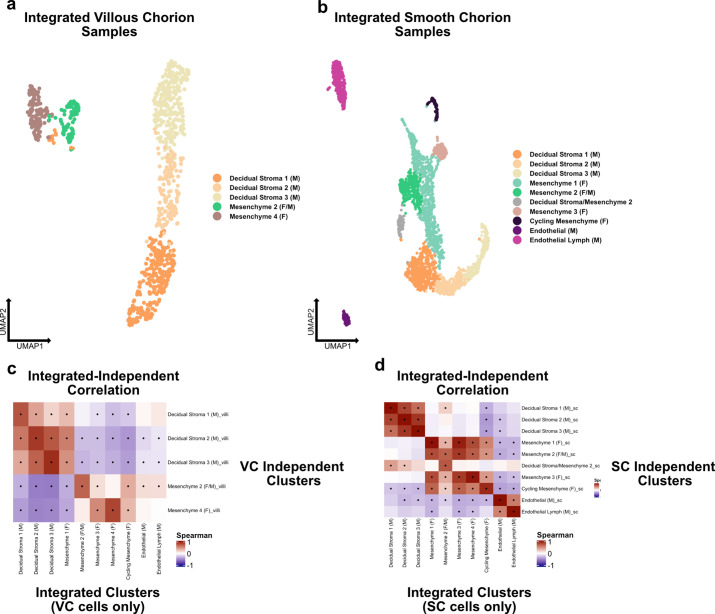
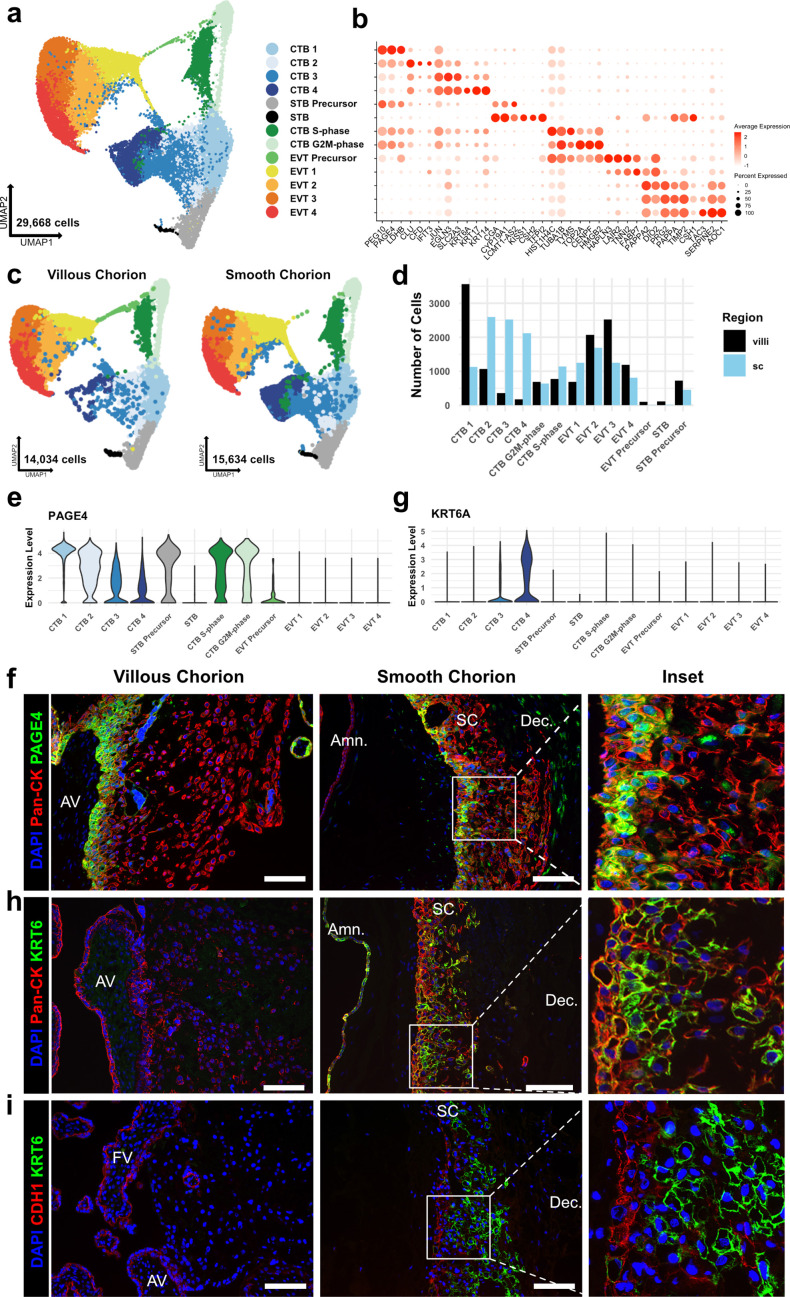
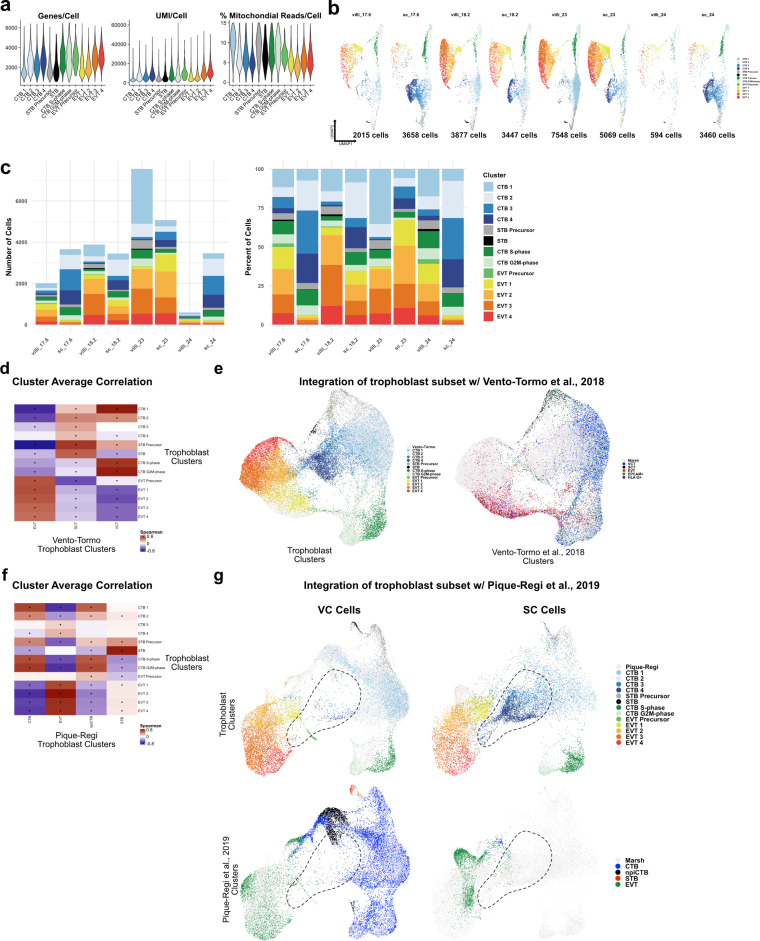
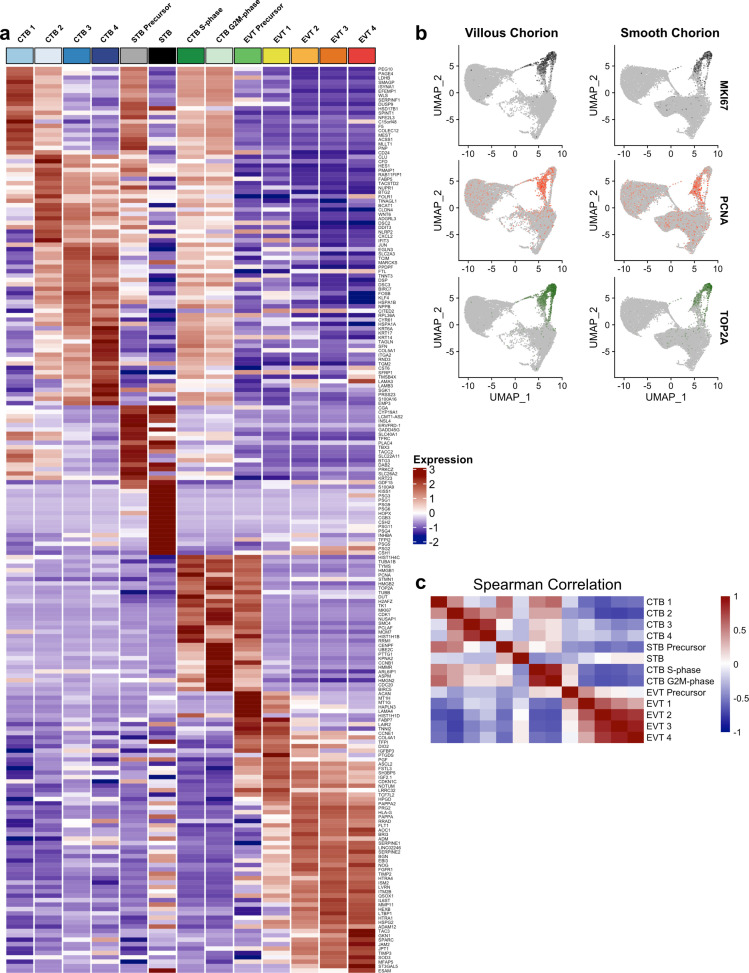
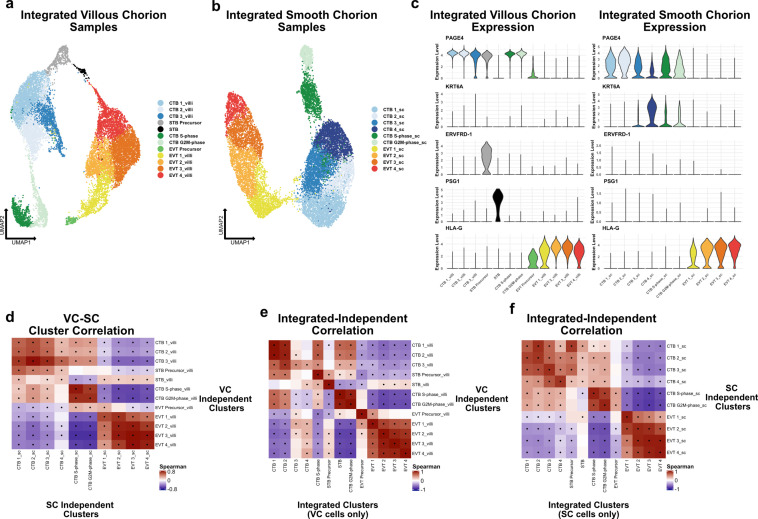
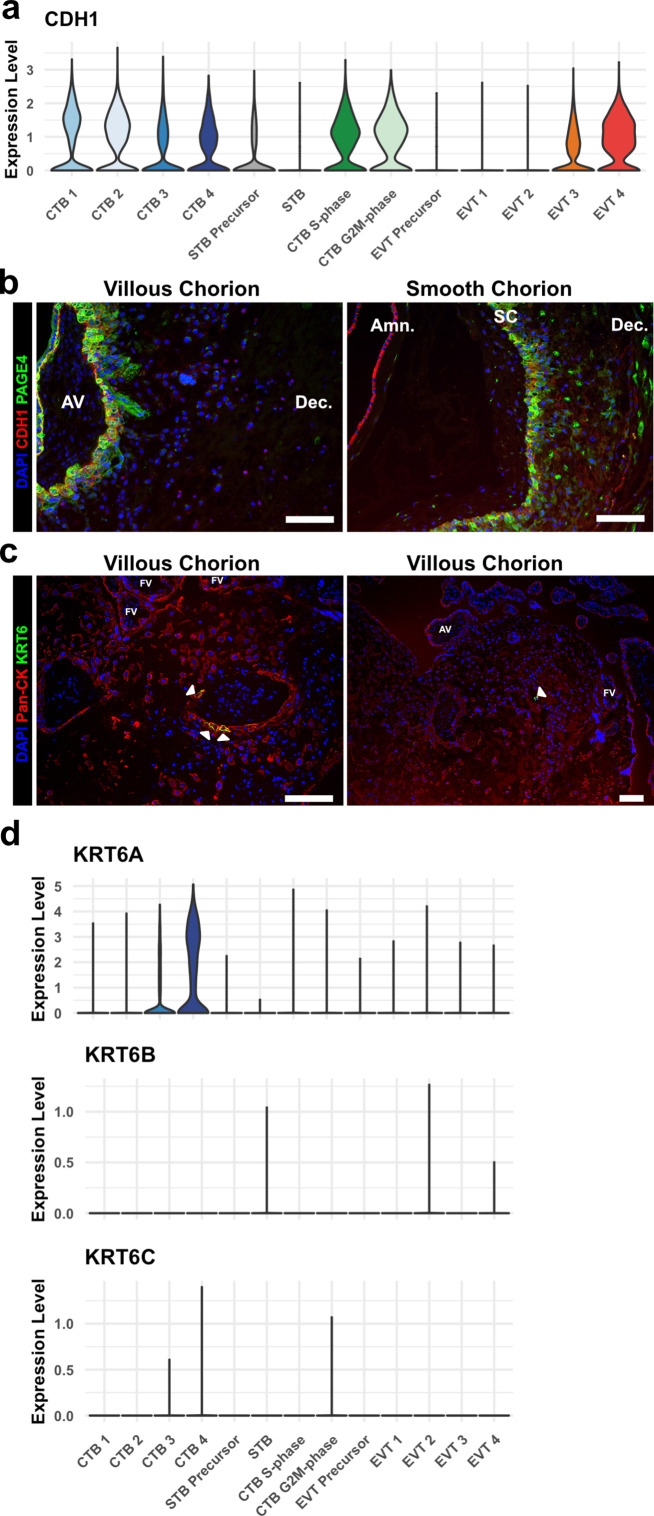
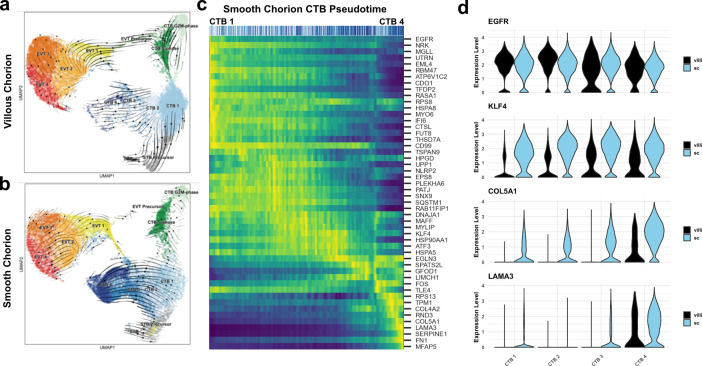
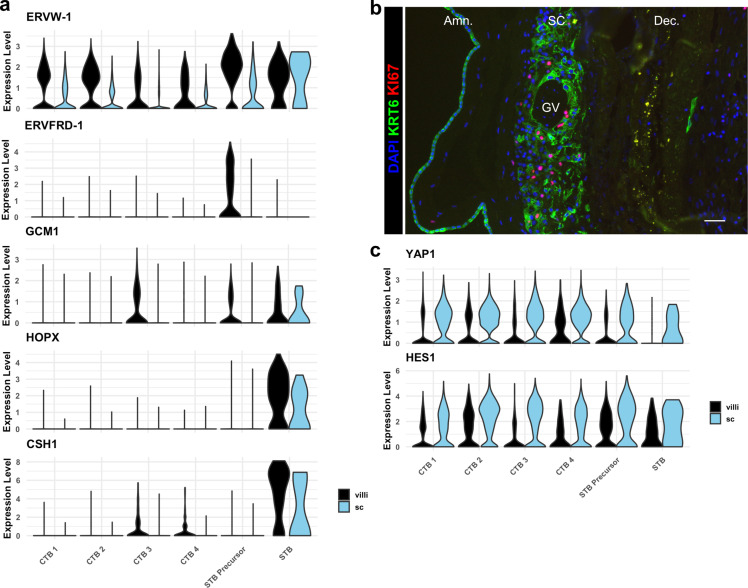
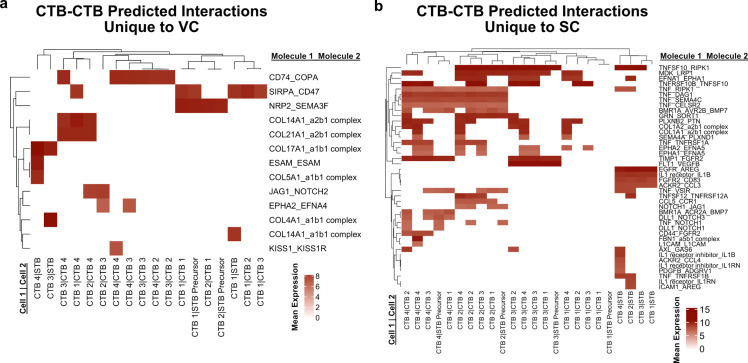
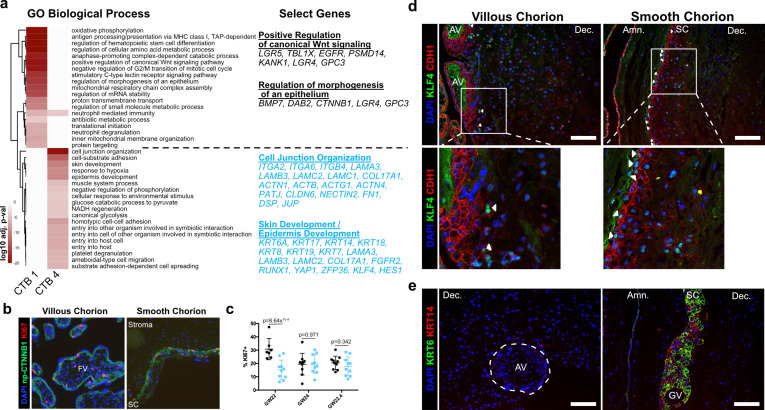
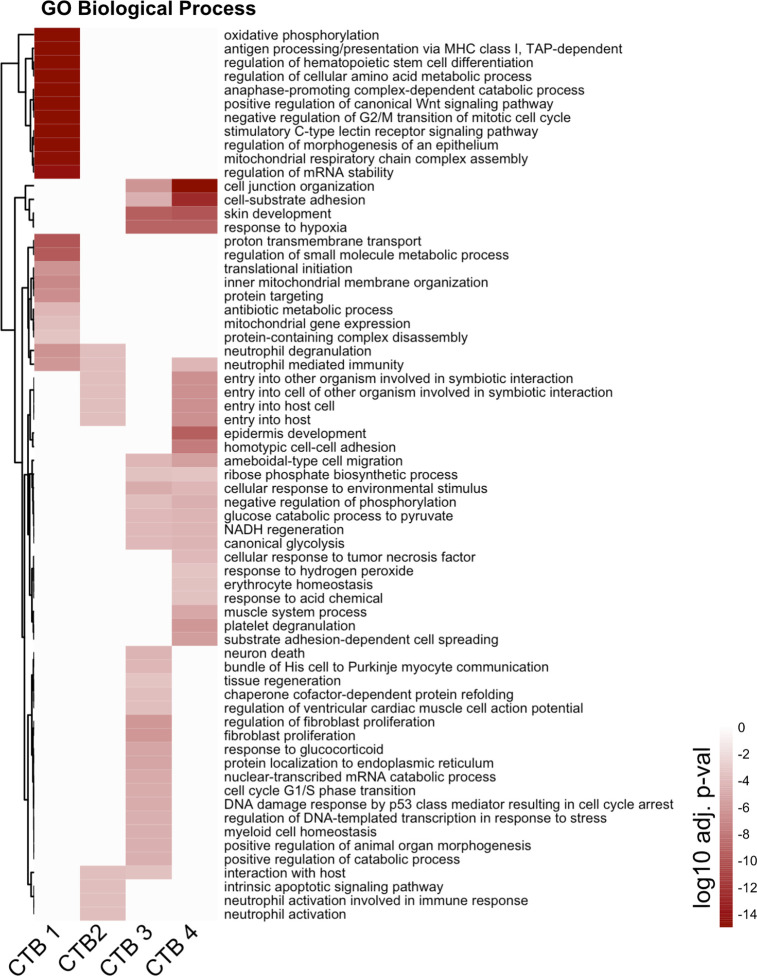
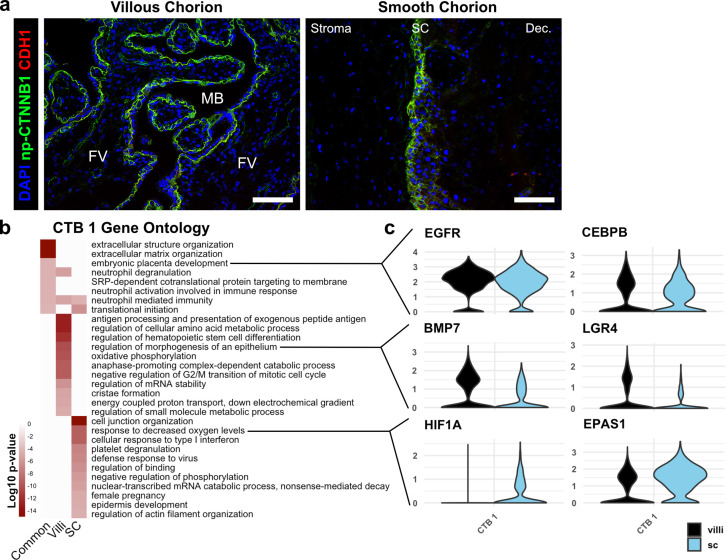
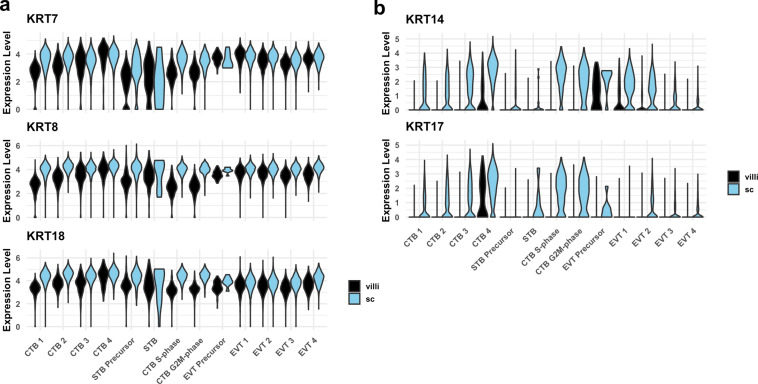
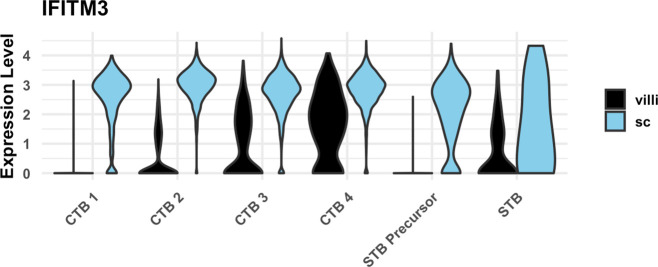
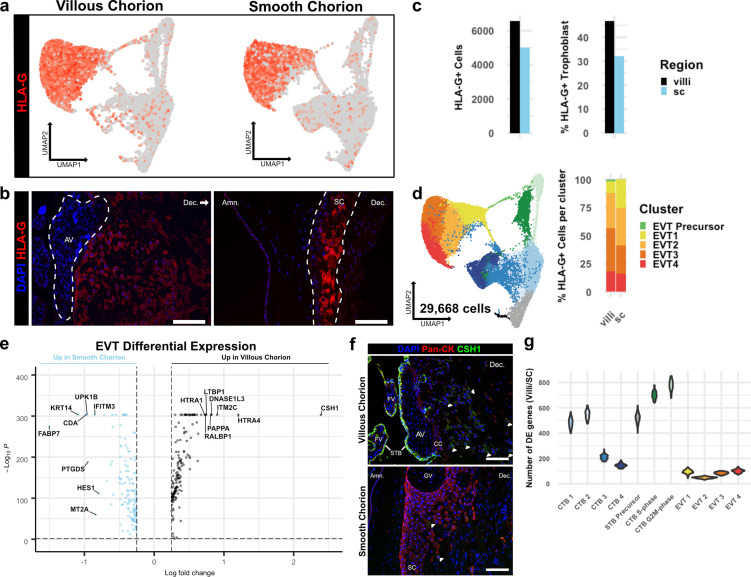
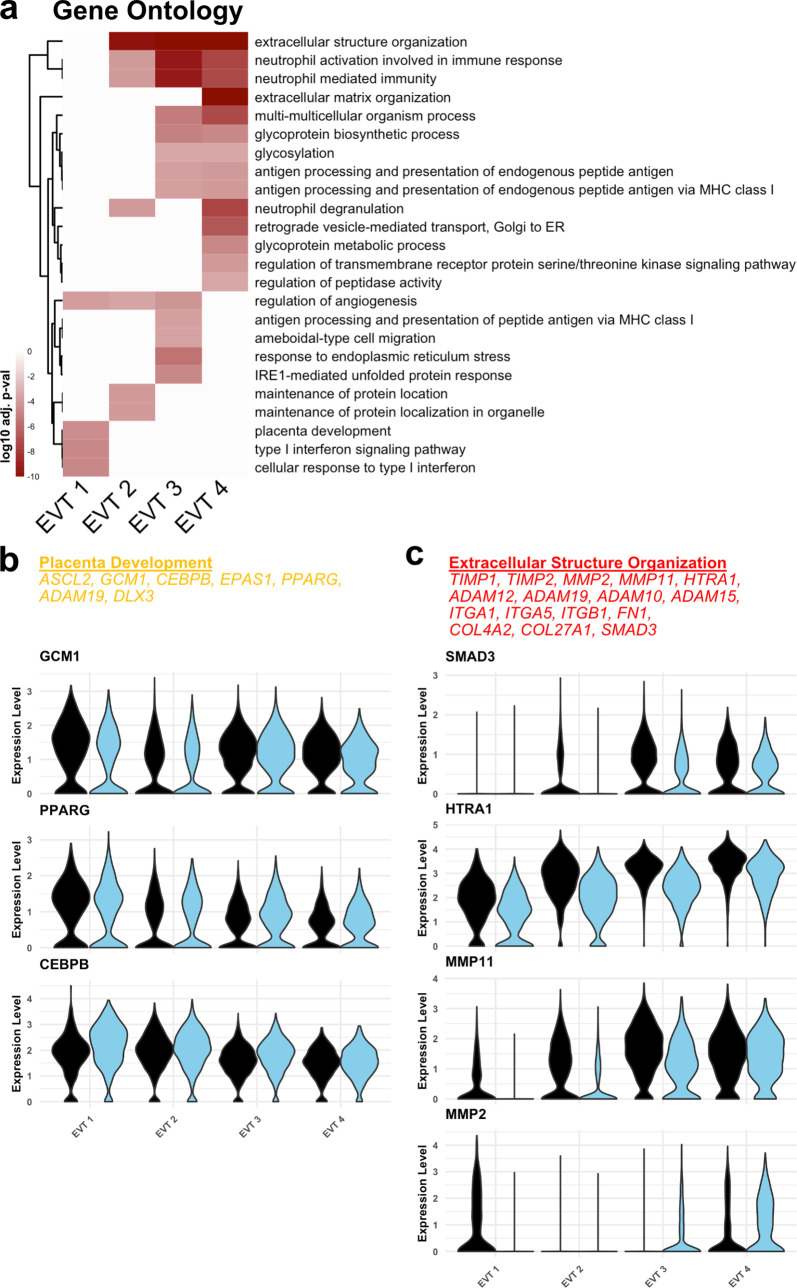
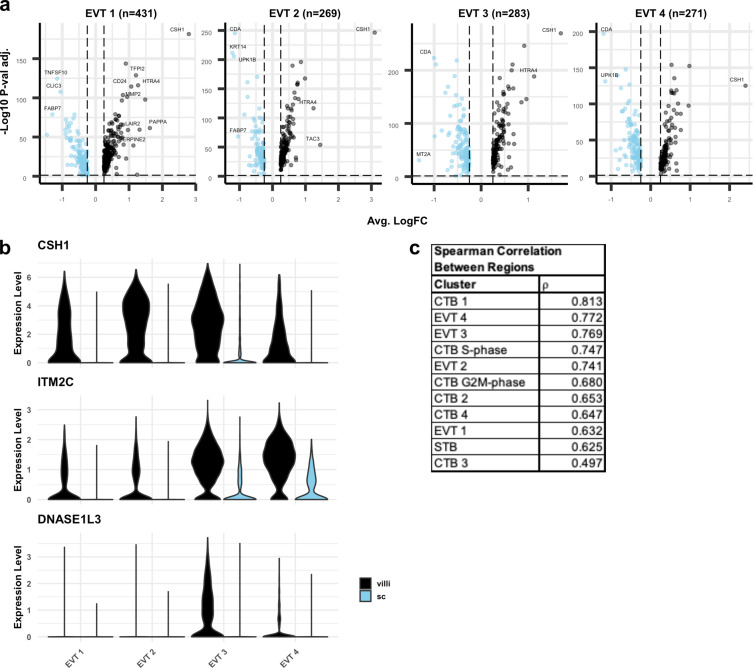
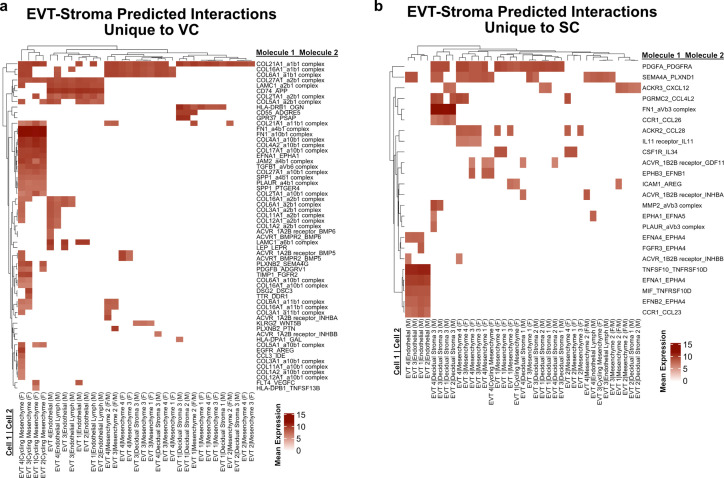
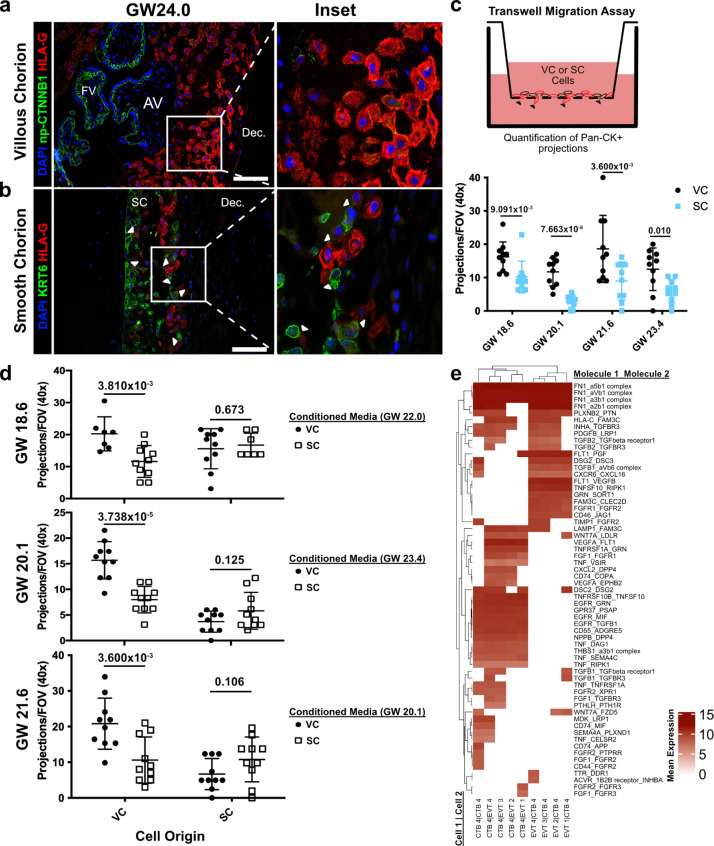
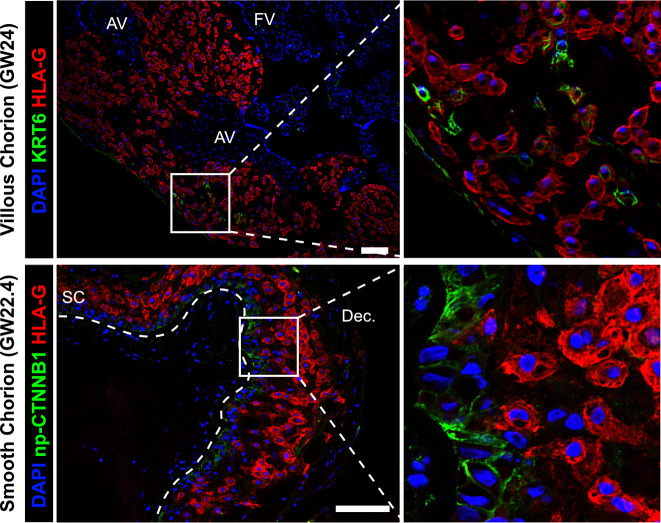
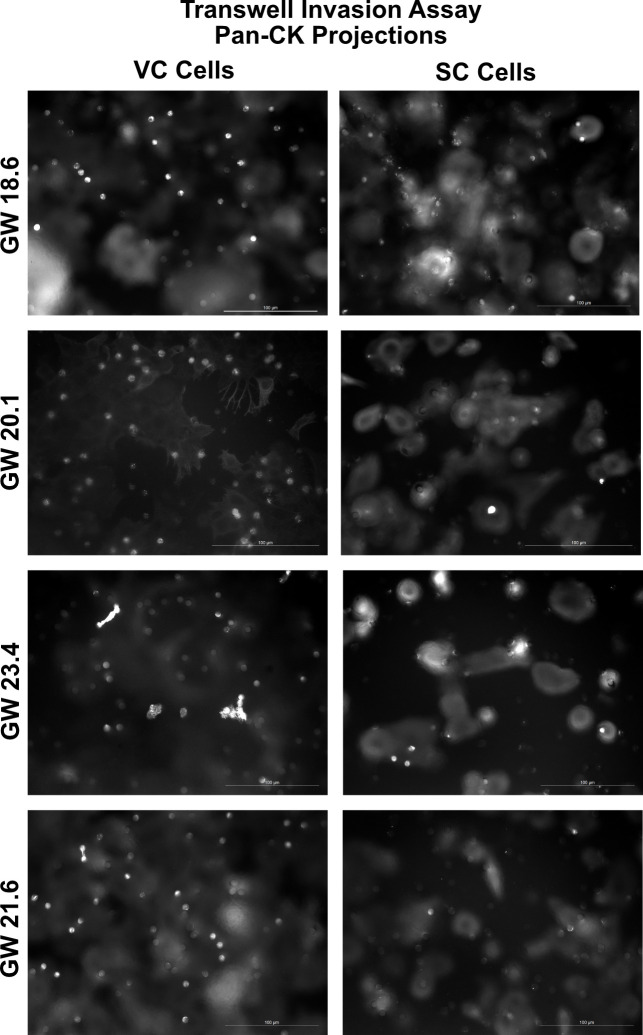
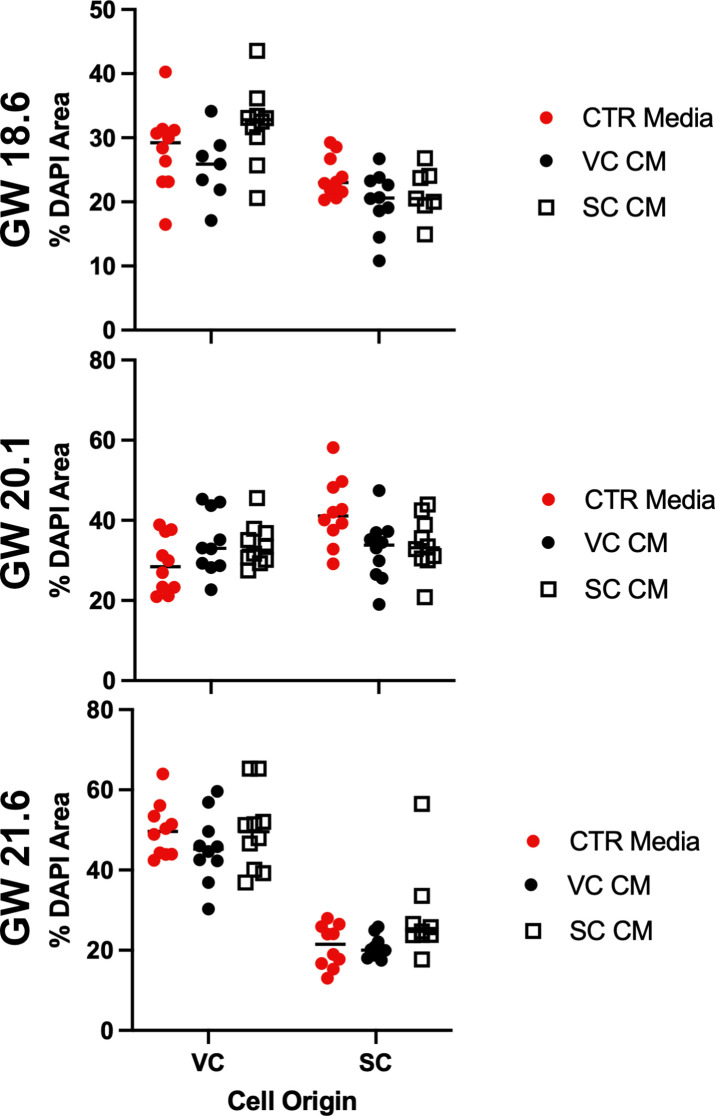
The human placenta contains two specialized regions: the villous chorion where gases and nutrients are exchanged between maternal and fetal blood, and the smooth chorion (SC) which surrounds more than 70% of the developing fetus but whose cellular composition and function is poorly understood. Here, we use single cell RNA-sequencing to compare the cell types and molecular programs between these two regions in the second trimester human placenta. Each region consists of progenitor cytotrophoblasts (CTBs) and extravillous trophoblasts (EVTs) with similar gene expression programs. While CTBs in the villous chorion differentiate into syncytiotrophoblasts, they take an alternative trajectory in the SC producing a previously unknown CTB population which we term SC-specific CTBs (SC-CTBs). Marked by expression of region-specific cytokeratins, the SC-CTBs form a stratified epithelium above a basal layer of progenitor CTBs. They express epidermal and metabolic transcriptional programs consistent with a primary role in defense against physical stress and pathogens. Additionally, we show that SC-CTBs closely associate with EVTs and secrete factors that inhibit the migration of the EVTs. This restriction of EVT migration is in striking contrast to the villous region where EVTs migrate away from the chorion and invade deeply into the decidua. Together, these findings greatly expand our understanding of CTB differentiation in these distinct regions of the human placenta. This knowledge has broad implications for studies of the development, functions, and diseases of the human placenta.
绒毛膜,其中母体和胎儿血液之间进行气体和营养物质的交换;以及平滑绒毛膜(SC),其围绕着发育中胎儿的 70%以上,但细胞组成和功能尚未被充分理解。在这里,我们使用单细胞 RNA 测序来比较这两个区域在人类胎盘中的细胞类型和分子程序。每个区域都包含祖细胞滋养层细胞(CTBs)和绒毛外滋养层细胞(EVTs),它们具有相似的基因表达程序。虽然绒毛膜中的 CTB 分化为合胞滋养层细胞,但它们在 SC 中采取了另一种轨迹,产生了一个以前未知的 CTB 群体,我们称之为 SC 特异性 CTB(SC-CTB)。SC-CTB 以表达区域特异性细胞角蛋白为特征,在上皮细胞基底层上方形成一个分层的上皮细胞层。它们表达表皮和代谢转录程序,与防御物理应激和病原体的主要作用一致。此外,我们还表明,SC-CTB 与 EVT 密切相关,并分泌抑制 EVT 迁移的因子。这种对 EVT 迁移的限制与绒毛膜区域形成鲜明对比,在绒毛膜区域,EVT 从绒毛膜迁移并深入侵入蜕膜。总之,这些发现极大地扩展了我们对这些人类胎盘不同区域 CTB 分化的理解。这一知识对于研究人类胎盘的发育、功能和疾病具有广泛的意义。