Mendoza-Hoffmann Francisco, Zarco-Zavala Mariel, Ortega Raquel, Celis-Sandoval Heliodoro, Torres-Larios Alfredo, García-Trejo José J
Facultad de Ciencias Químicas e Ingeniería, Universidad Autónoma de Baja California (UABC)-Campus Tijuana, Tijuana C.P. 22390, Baja California, Mexico.
Departamento de Biología, Facultad de Química, Ciudad Universitaria, Universidad Nacional Autónoma de México (U.N.A.M.), Ciudad de Mexico C.P. 04510, Coyoacan, Mexico.
Microorganisms. 2022 Jul 7;10(7):1372. doi: 10.3390/microorganisms10071372.
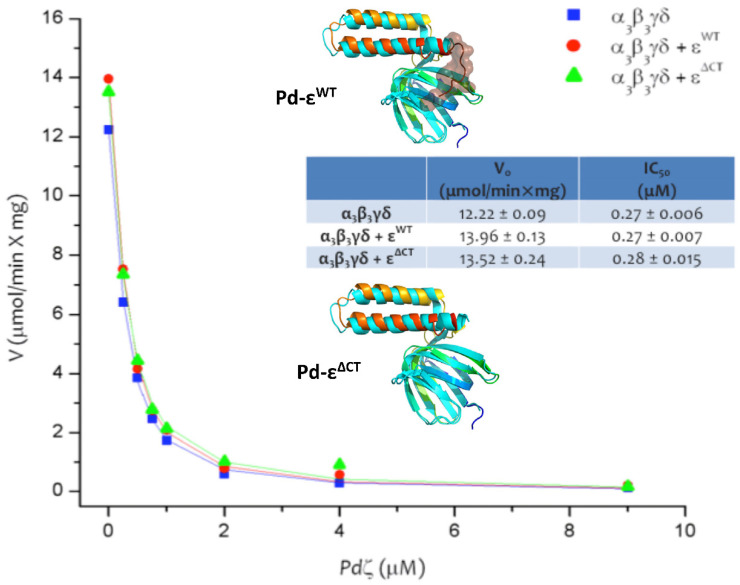
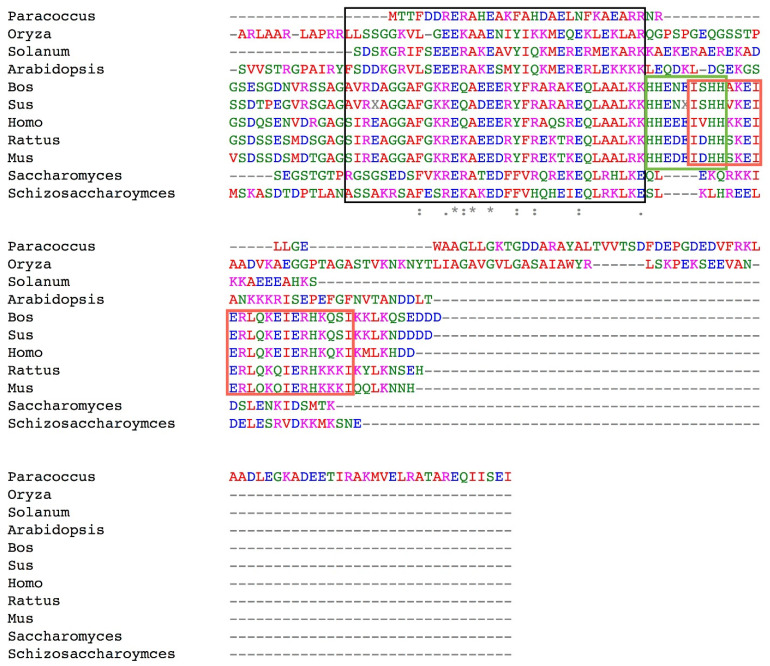
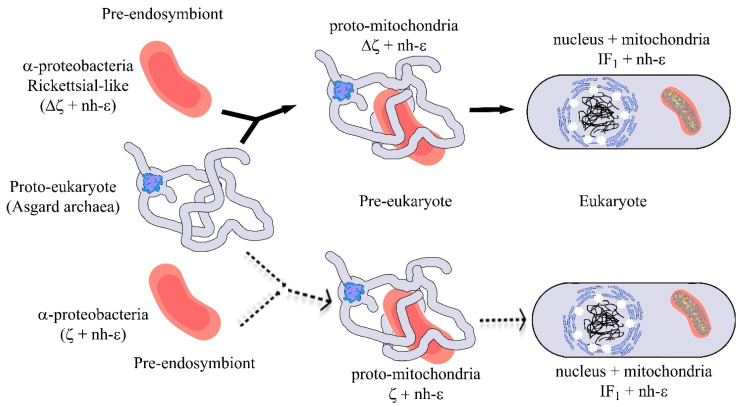
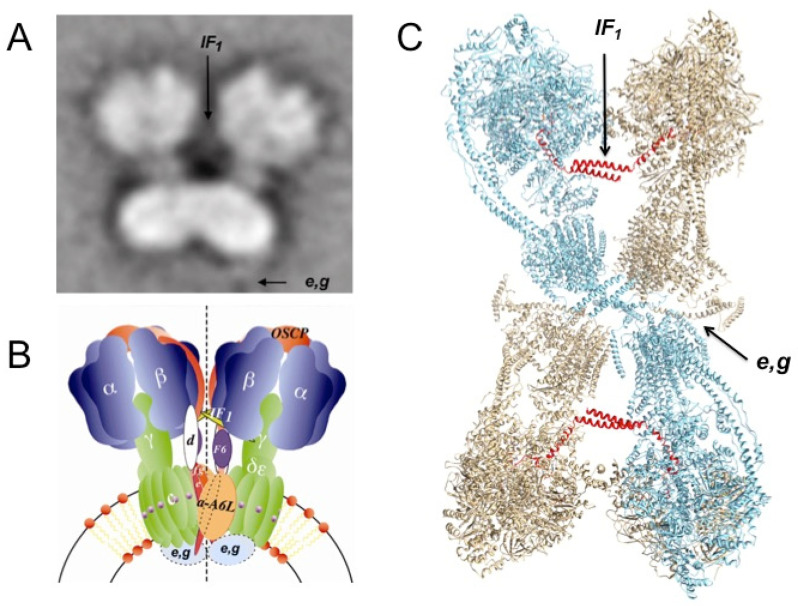
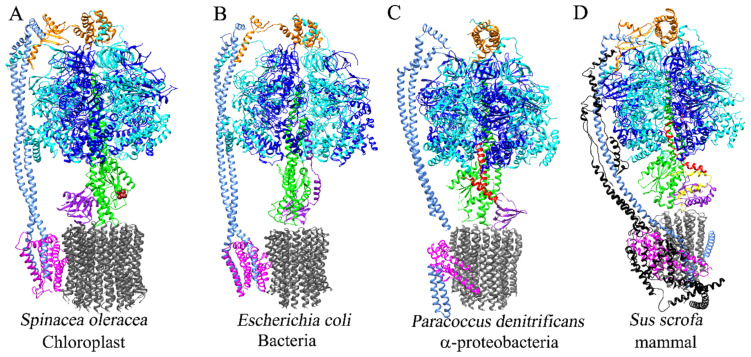
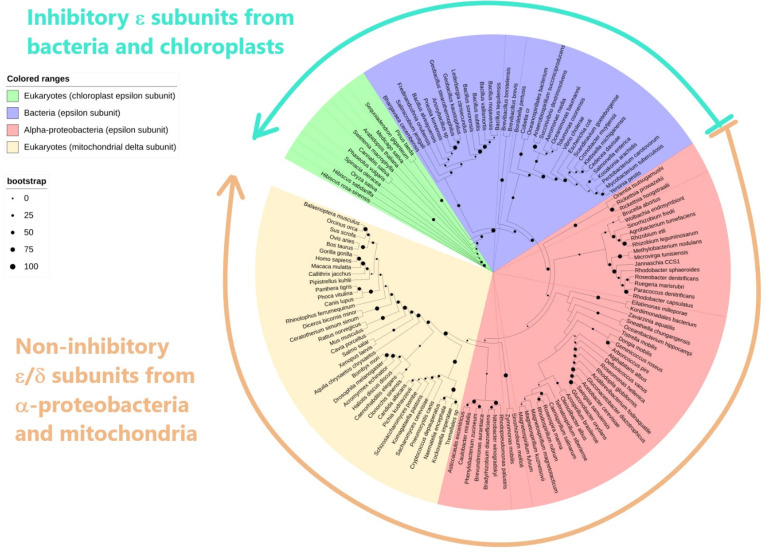
The F1FO-ATP synthase nanomotor synthesizes >90% of the cellular ATP of almost all living beings by rotating in the “forward” direction, but it can also consume the same ATP pools by rotating in “reverse.” To prevent futile F1FO-ATPase activity, several different inhibitory proteins or domains in bacteria (ε and ζ subunits), mitochondria (IF1), and chloroplasts (ε and γ disulfide) emerged to block the F1FO-ATPase activity selectively. In this study, we analyze how these F1FO-ATPase inhibitory proteins have evolved. The phylogeny of the α-proteobacterial ε showed that it diverged in its C-terminal side, thus losing both the inhibitory function and the ATP-binding/sensor motif that controls this inhibition. The losses of inhibitory function and the ATP-binding site correlate with an evolutionary divergence of non-inhibitory α-proteobacterial ε and mitochondrial δ subunits from inhibitory bacterial and chloroplastidic ε subunits. Here, we confirm the lack of inhibitory function of wild-type and C-terminal truncated ε subunits of P. denitrificans. Taken together, the data show that ζ evolved to replace ε as the primary inhibitor of the F1FO-ATPase of free-living α-proteobacteria. However, the ζ inhibitory function was also partially lost in some symbiotic α-proteobacteria and totally lost in some strictly parasitic α-proteobacteria such as the Rickettsiales order. Finally, we found that ζ and IF1 likely evolved independently via convergent evolution before and after the endosymbiotic origin mitochondria, respectively. This led us to propose the ε and ζ subunits as tracer genes of the pre-endosymbiont that evolved into the actual mitochondria.
F1FO - ATP合酶纳米马达通过“正向”旋转合成几乎所有生物细胞中90%以上的ATP,但它也可以通过“反向”旋转消耗相同的ATP库。为了防止徒劳的F1FO - ATP酶活性,细菌(ε和ζ亚基)、线粒体(IF1)和叶绿体(ε和γ二硫键)中出现了几种不同的抑制蛋白或结构域,以选择性地阻断F1FO - ATP酶活性。在本研究中,我们分析了这些F1FO - ATP酶抑制蛋白是如何进化的。α - 变形菌属ε的系统发育表明,它在C端发生了分化,从而失去了抑制功能以及控制这种抑制的ATP结合/传感基序。抑制功能和ATP结合位点的丧失与非抑制性α - 变形菌属ε和线粒体δ亚基与抑制性细菌和叶绿体ε亚基的进化分化相关。在这里,我们证实了反硝化假单胞菌野生型和C端截短的ε亚基缺乏抑制功能。综上所述,数据表明ζ进化为取代ε,成为自由生活的α - 变形菌F1FO - ATP酶的主要抑制剂。然而,ζ的抑制功能在一些共生α - 变形菌中也部分丧失,在一些严格寄生的α - 变形菌如立克次氏体目中完全丧失。最后,我们发现ζ和IF1可能分别在内共生起源线粒体之前和之后通过趋同进化独立进化。这使我们提出ε和ζ亚基作为进化为实际线粒体的内共生体前体的示踪基因。