Division of Molecular Pathology, Netherlands Cancer Institute, Amsterdam, The Netherlands.
Oncode Institute, Utrecht, The Netherlands.
Nature. 2022 Aug;608(7923):609-617. doi: 10.1038/s41586-022-05066-5. Epub 2022 Aug 10.
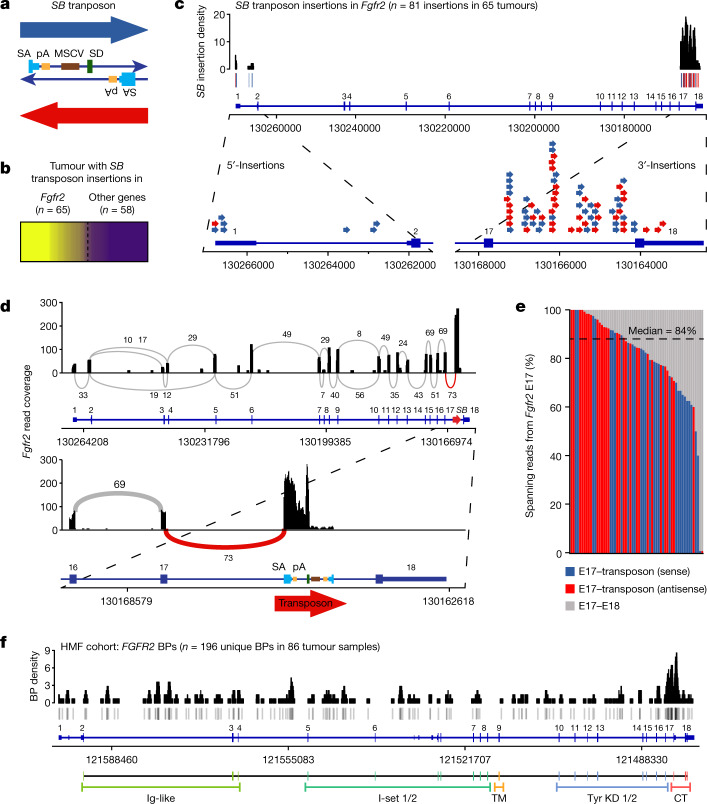
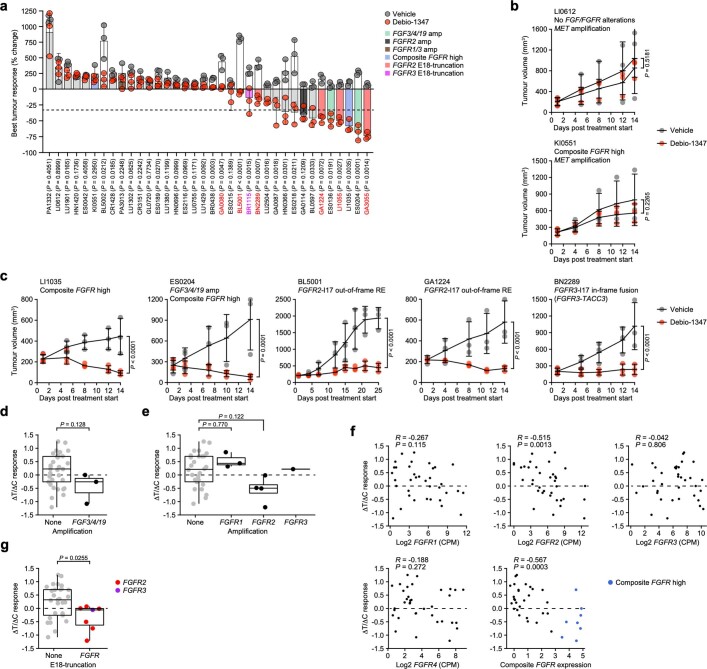
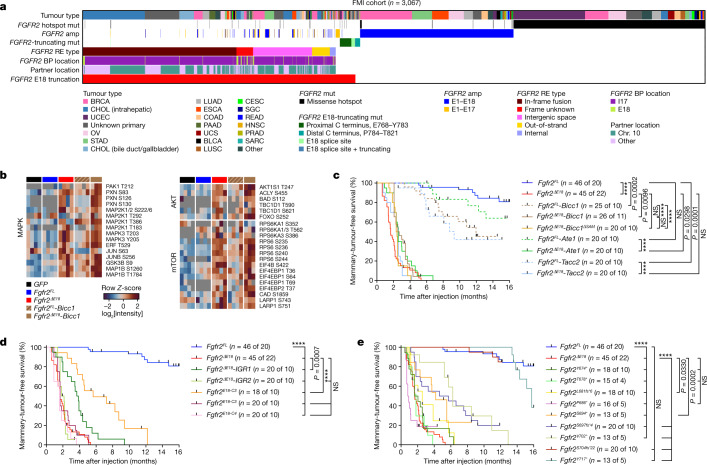
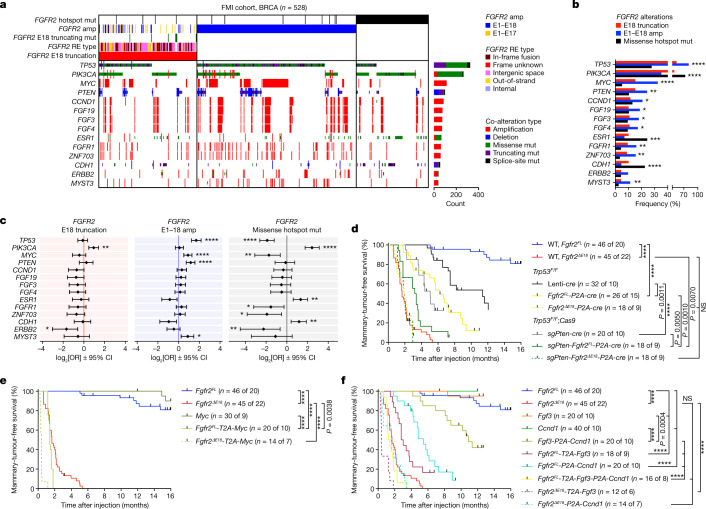
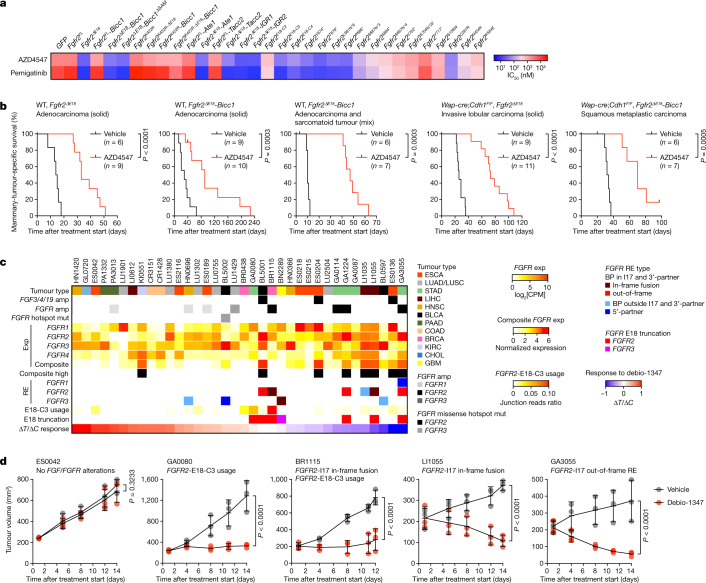
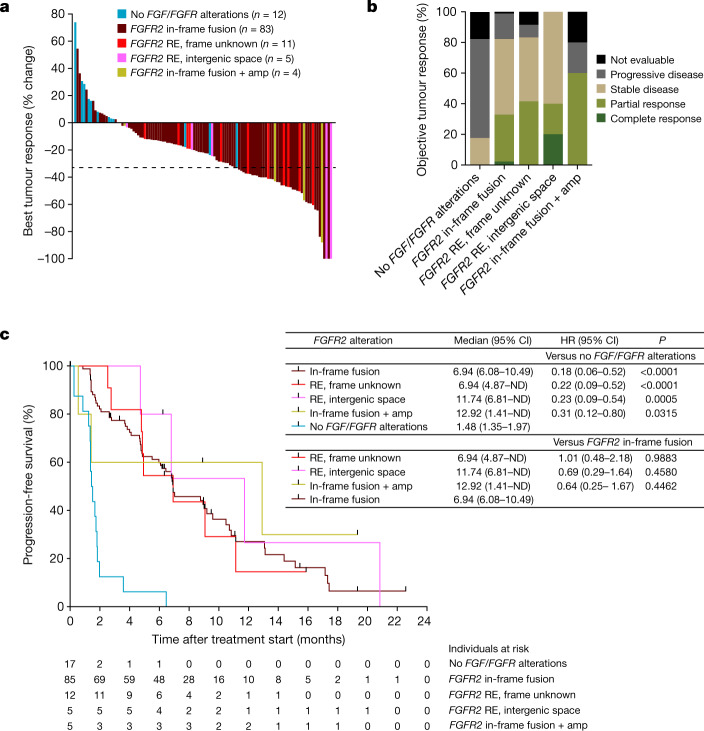
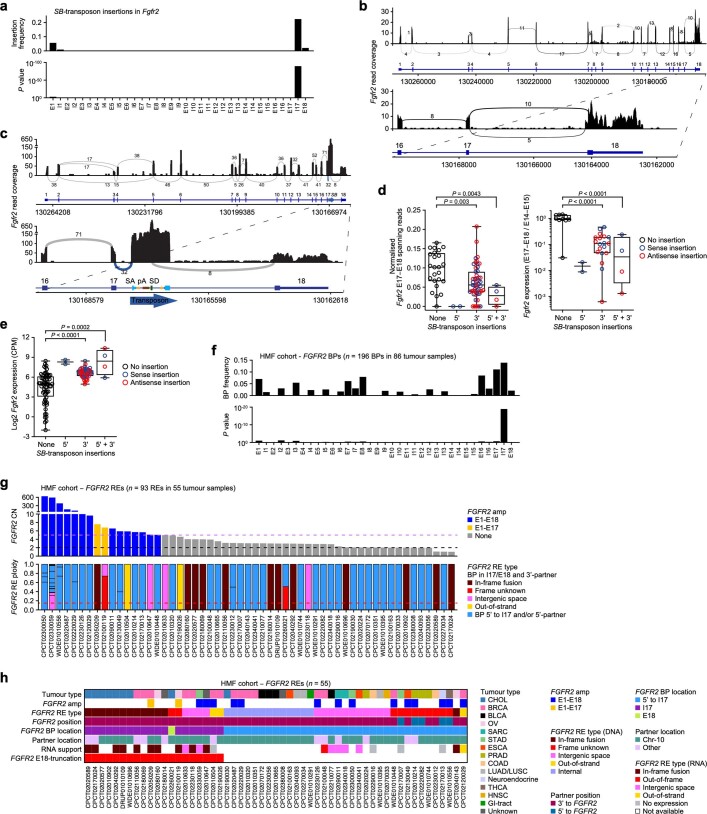
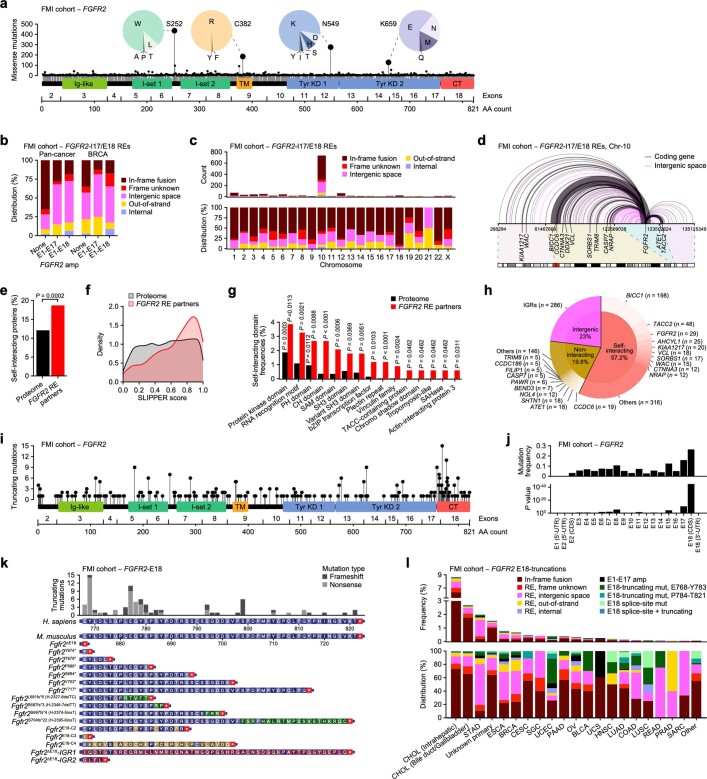
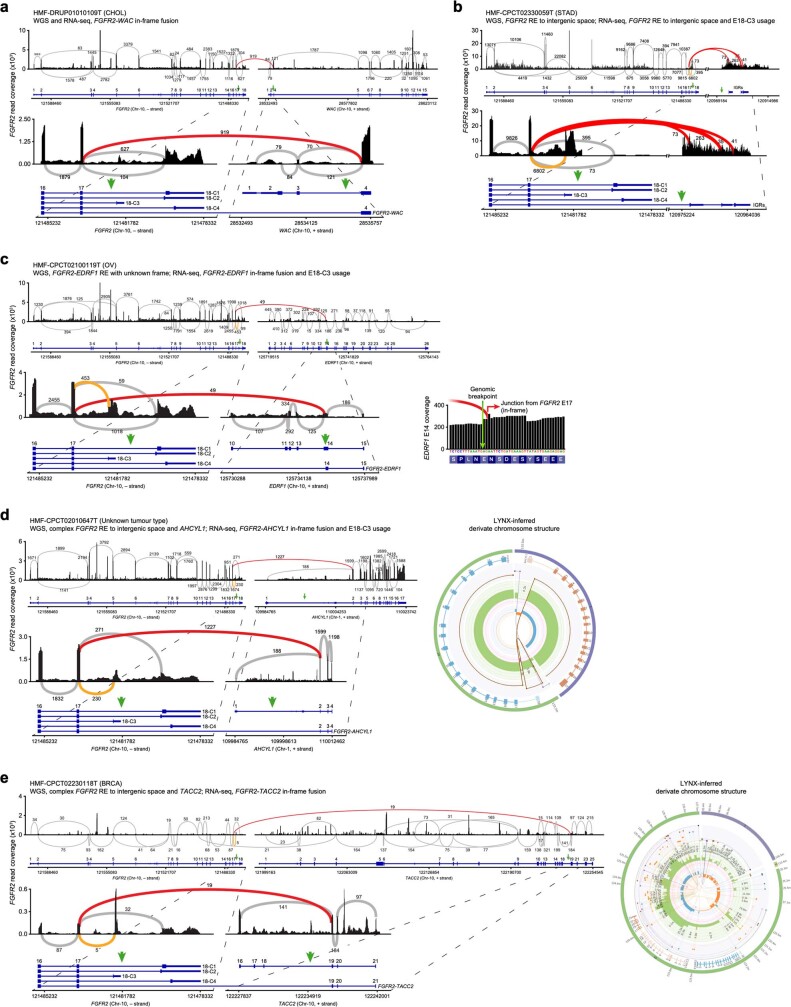
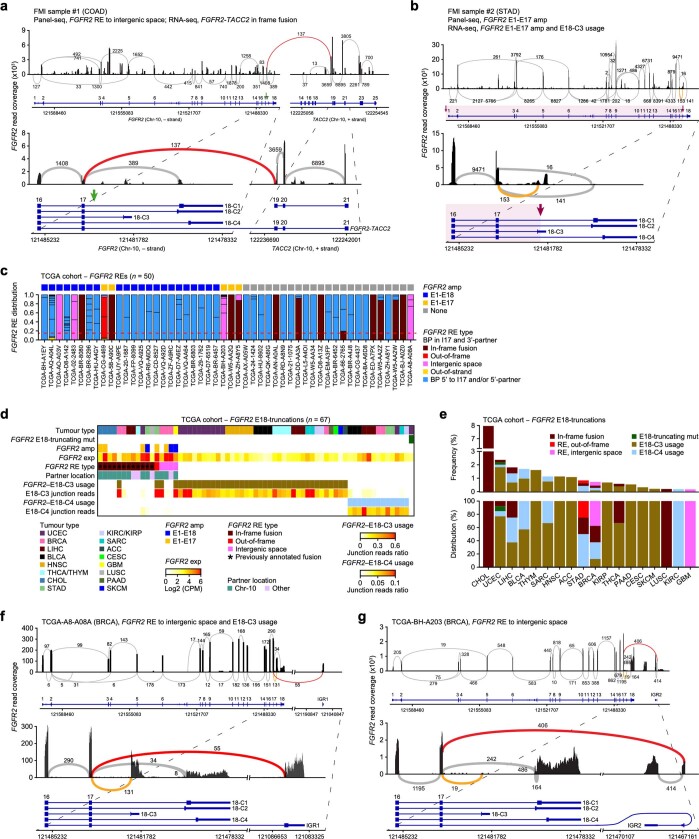
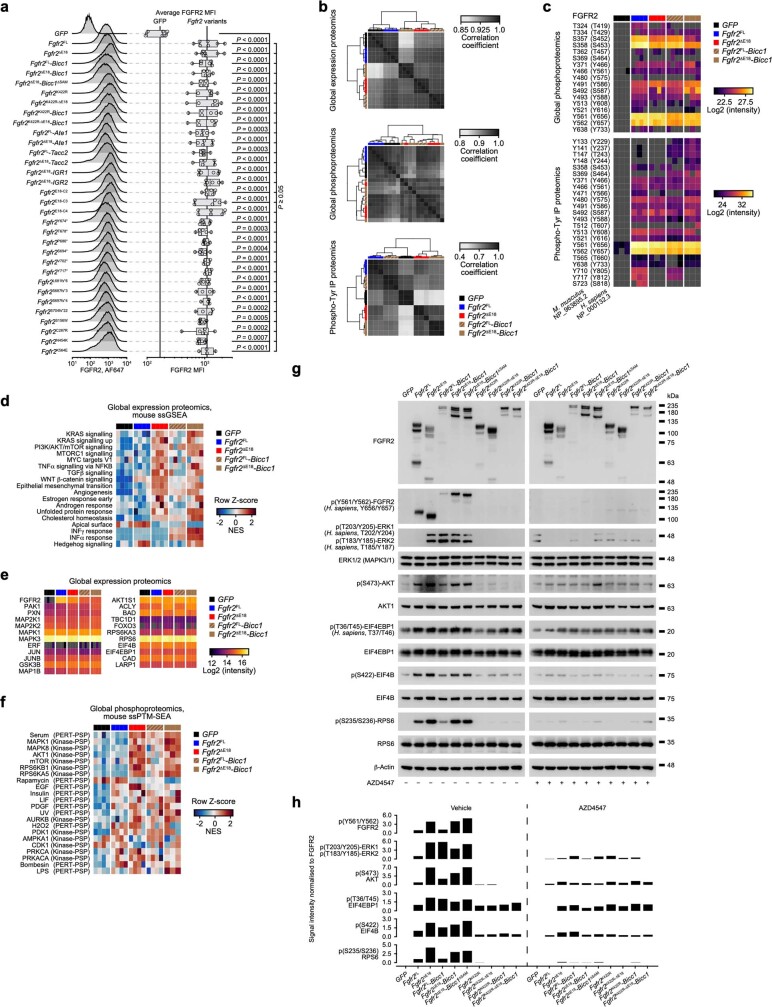
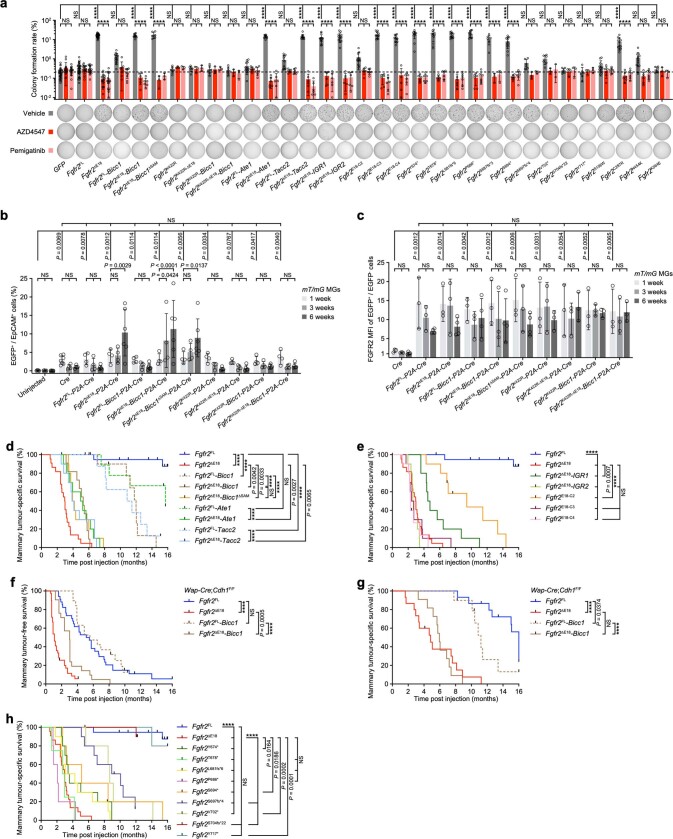
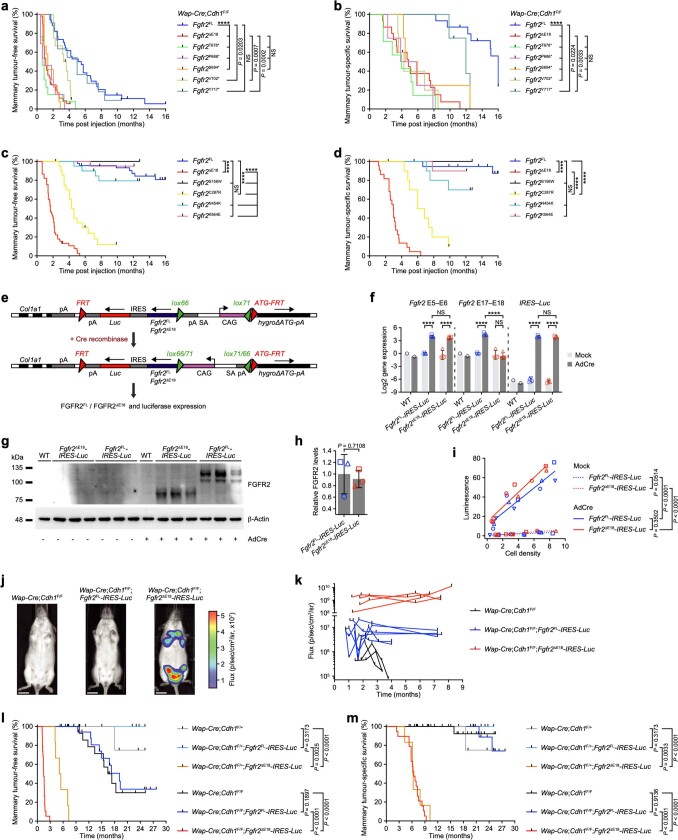
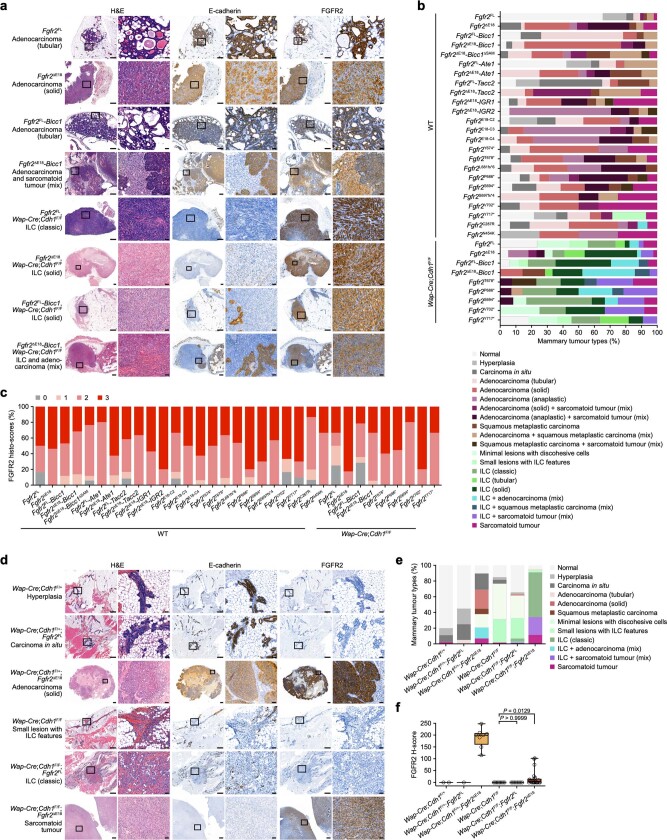
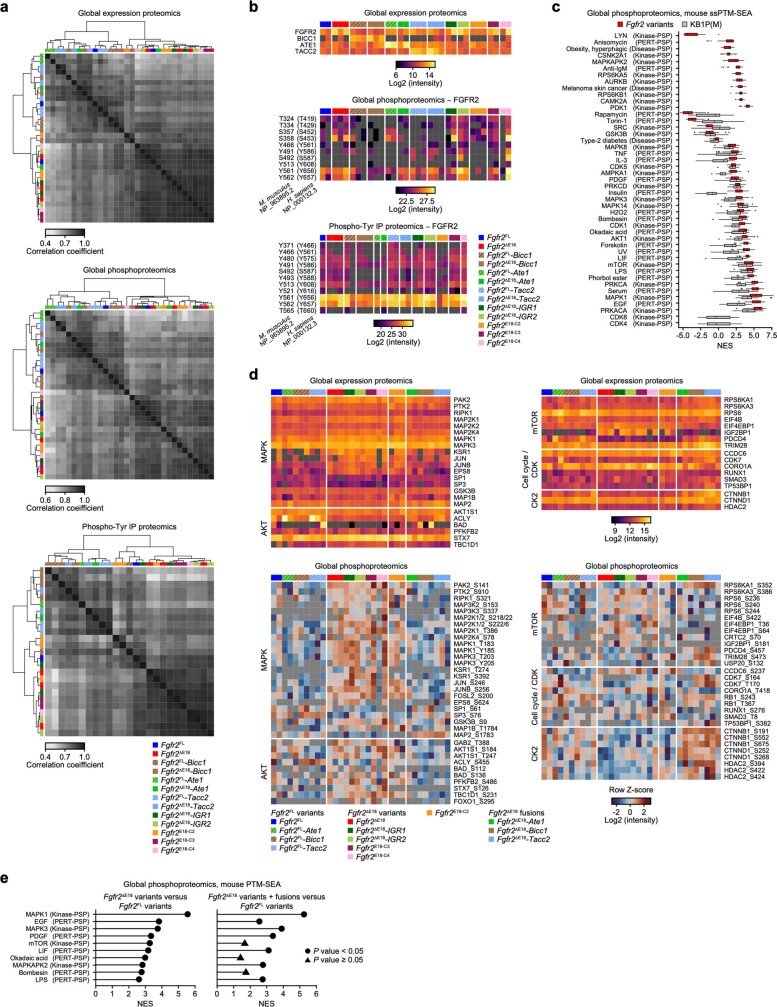
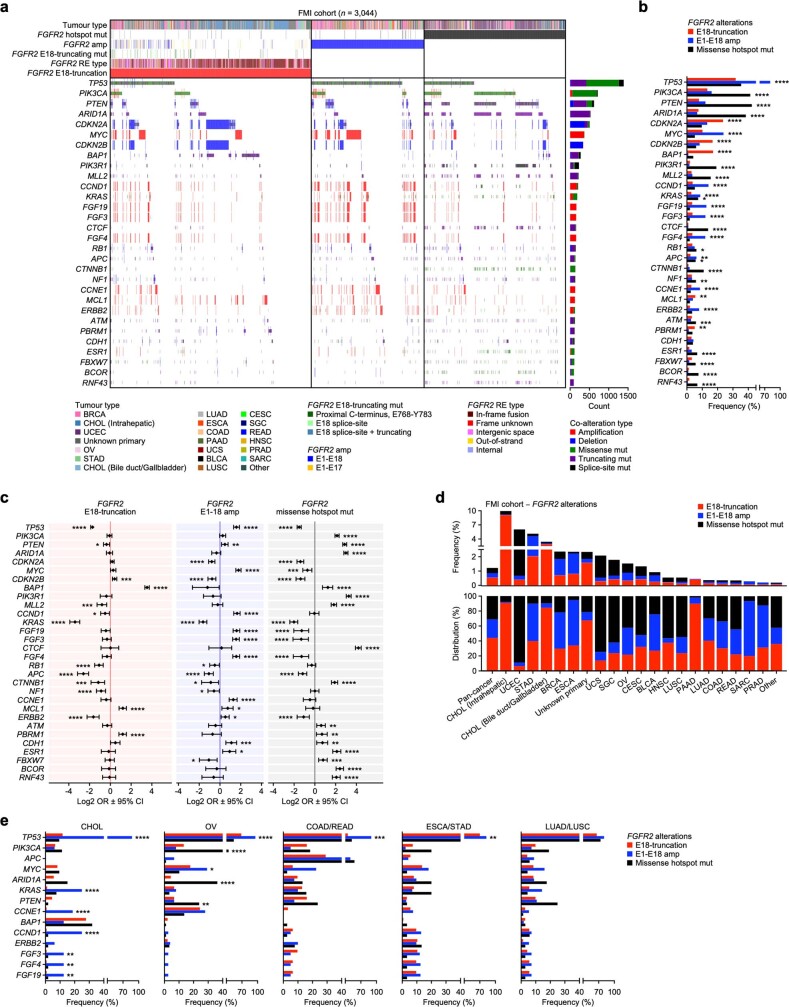
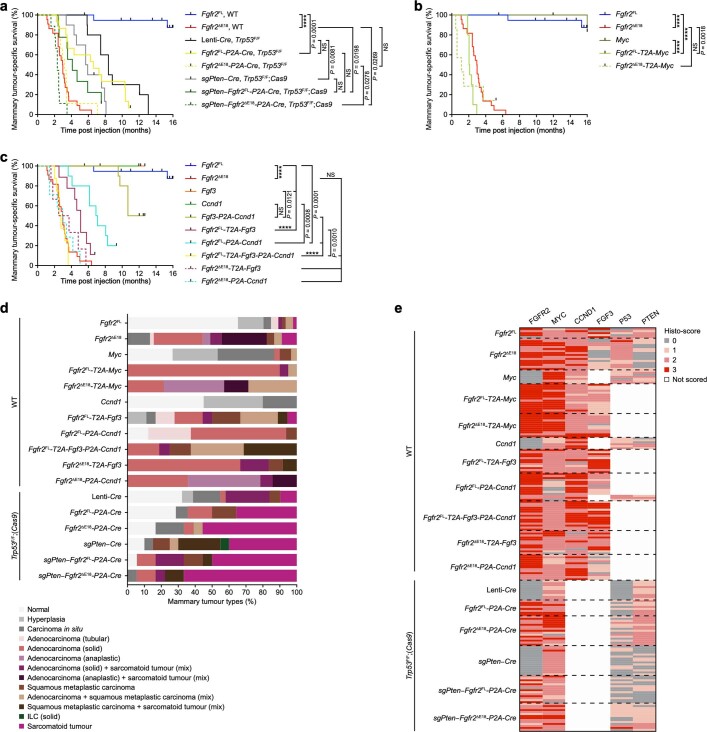
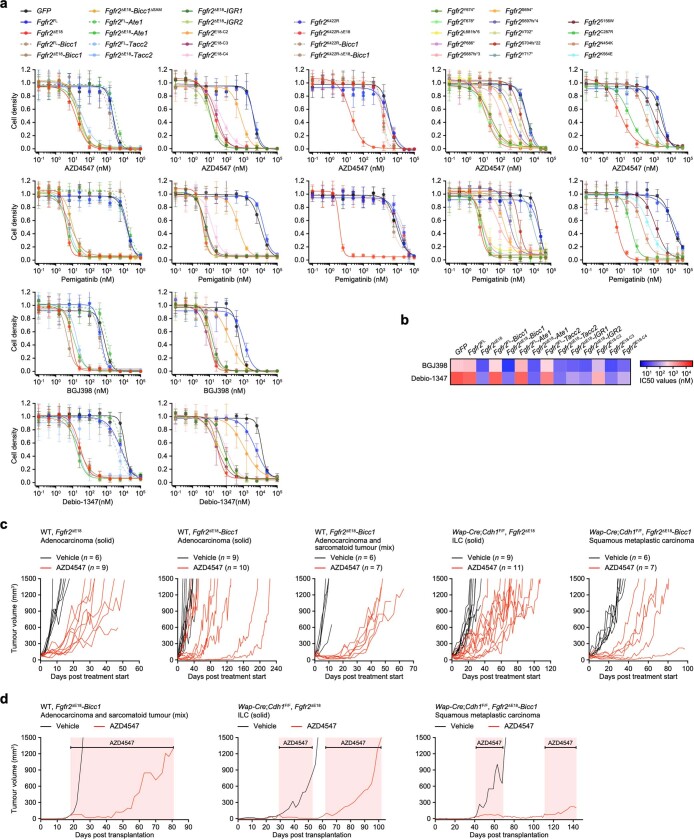
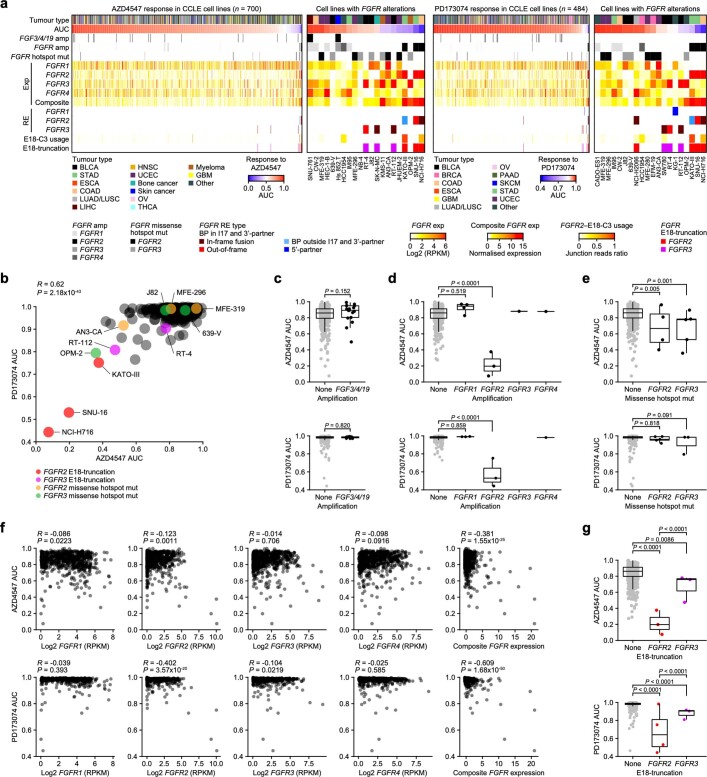
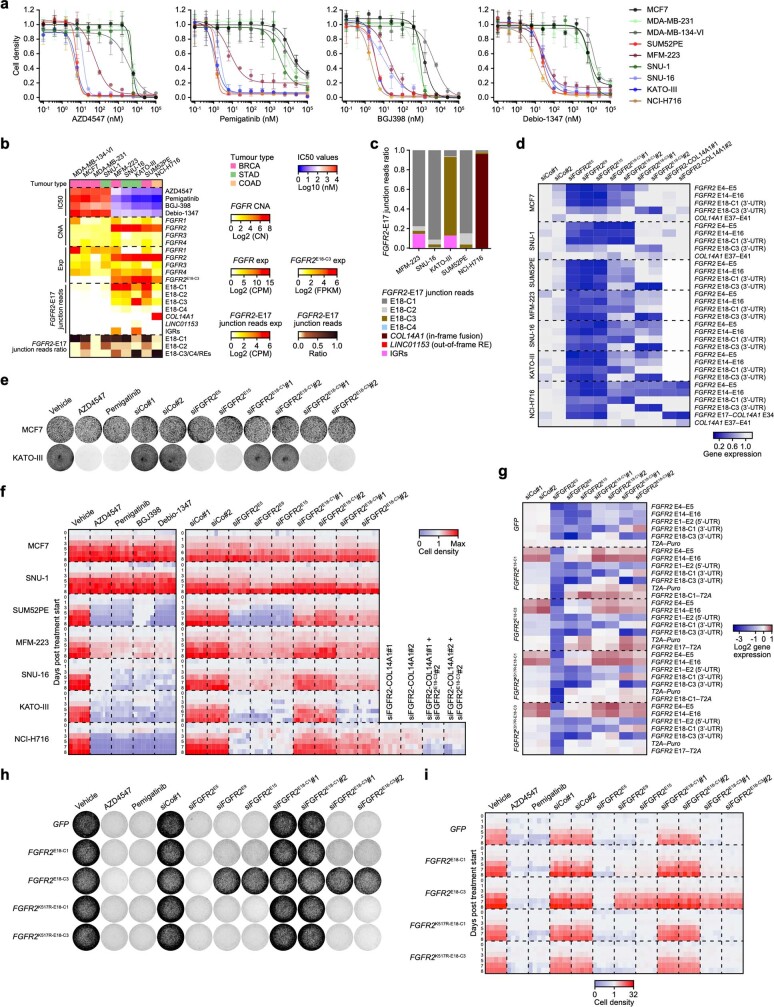
Somatic hotspot mutations and structural amplifications and fusions that affect fibroblast growth factor receptor 2 (encoded by FGFR2) occur in multiple types of cancer. However, clinical responses to FGFR inhibitors have remained variable, emphasizing the need to better understand which FGFR2 alterations are oncogenic and therapeutically targetable. Here we apply transposon-based screening and tumour modelling in mice, and find that the truncation of exon 18 (E18) of Fgfr2 is a potent driver mutation. Human oncogenomic datasets revealed a diverse set of FGFR2 alterations, including rearrangements, E1-E17 partial amplifications, and E18 nonsense and frameshift mutations, each causing the transcription of E18-truncated FGFR2 (FGFR2). Functional in vitro and in vivo examination of a compendium of FGFR2 and full-length variants pinpointed FGFR2-E18 truncation as single-driver alteration in cancer. By contrast, the oncogenic competence of FGFR2 full-length amplifications depended on a distinct landscape of cooperating driver genes. This suggests that genomic alterations that generate stable FGFR2 variants are actionable therapeutic targets, which we confirmed in preclinical mouse and human tumour models, and in a clinical trial. We propose that cancers containing any FGFR2 variant with a truncated E18 should be considered for FGFR-targeted therapies.
体细胞热点突变和结构扩增与融合影响成纤维细胞生长因子受体 2(由 FGFR2 编码),发生在多种类型的癌症中。然而,针对 FGFR 抑制剂的临床反应仍然各不相同,这强调了需要更好地了解哪些 FGFR2 改变是致癌的和可治疗靶向的。在这里,我们应用转座子为基础的筛选和肿瘤建模在小鼠中,并发现 FGFR2 外显子 18(E18)的截断是一个有效的驱动突变。人类致癌基因组数据集揭示了一系列不同的 FGFR2 改变,包括重排、E1-E17 部分扩增以及 E18 无义和移码突变,每个都导致 E18 截断的 FGFR2(FGFR2)转录。对 FGFR2 和全长变体的综合汇编的体外和体内功能检查将 FGFR2-E18 截断鉴定为癌症中的单一驱动改变。相比之下,全长 FGFR2 扩增的致癌能力取决于不同的合作驱动基因景观。这表明产生稳定 FGFR2 变体的基因组改变是可操作的治疗靶点,我们在临床前小鼠和人类肿瘤模型以及临床试验中得到了证实。我们建议将任何包含 E18 截断的 FGFR2 变体的癌症都应考虑进行 FGFR 靶向治疗。