Department of Pharmacology & Cancer Biology, Duke University Medical Center, Durham, United States.
Department of Pathology, Duke University Medical Center, Durham, United States.
Elife. 2022 Sep 7;11:e75715. doi: 10.7554/eLife.75715.
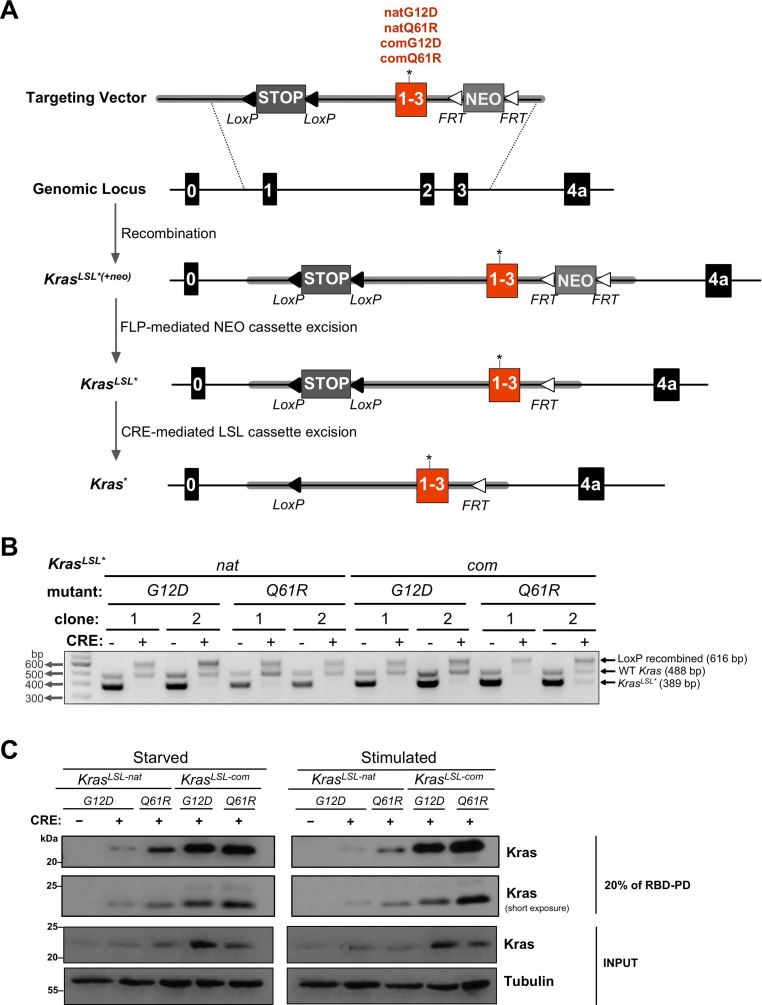
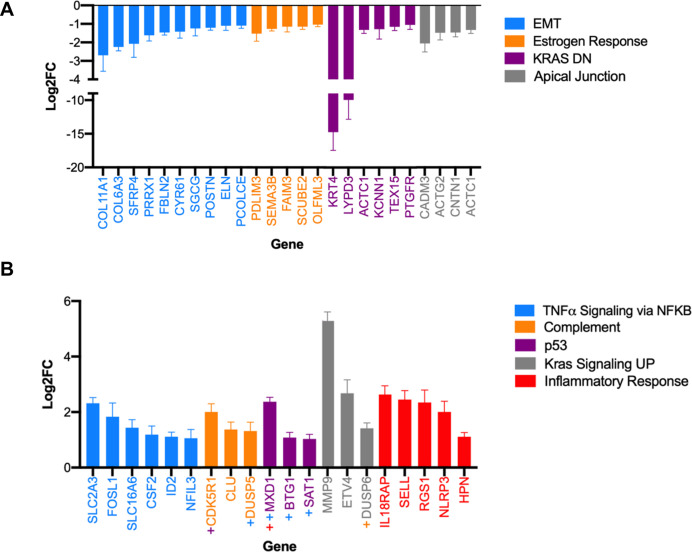
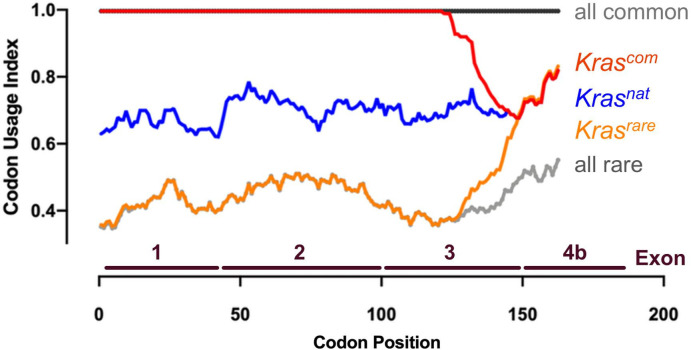
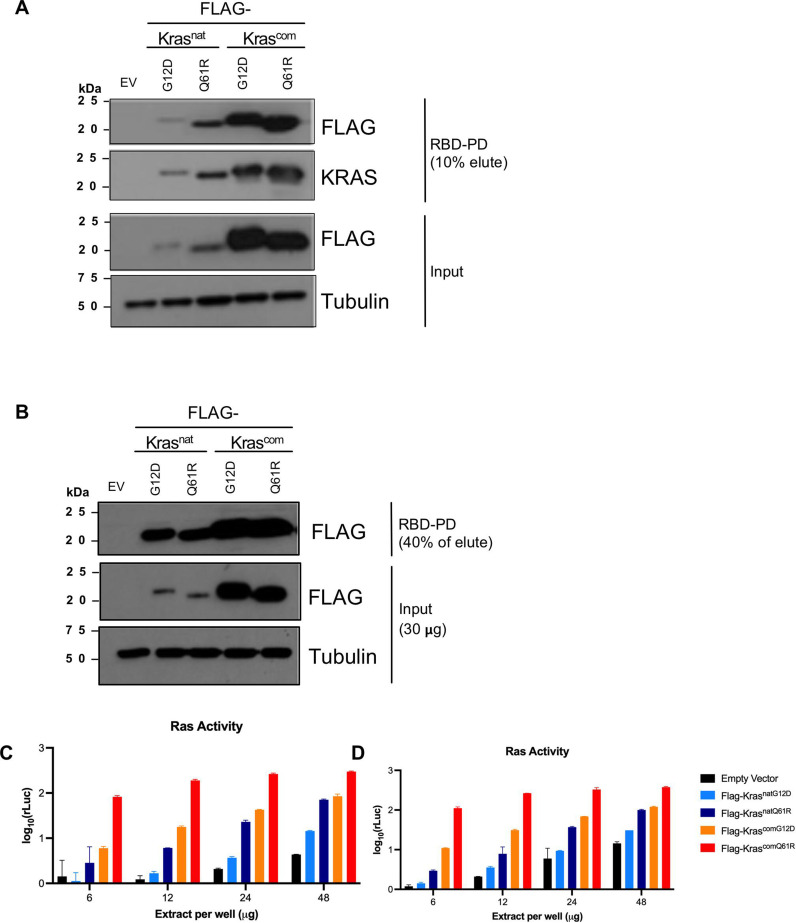
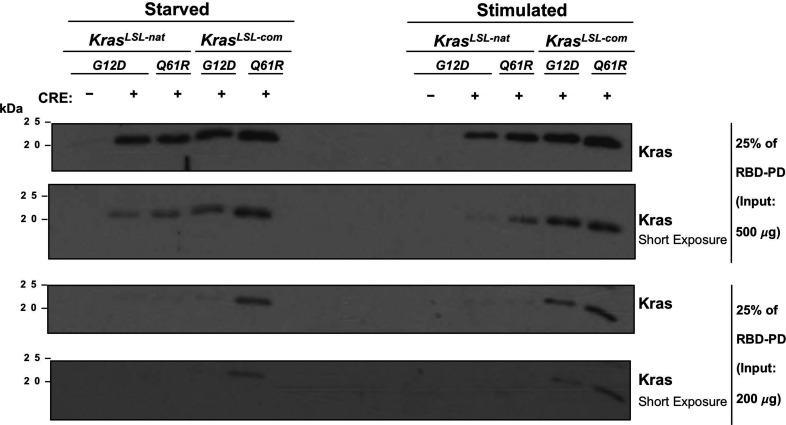
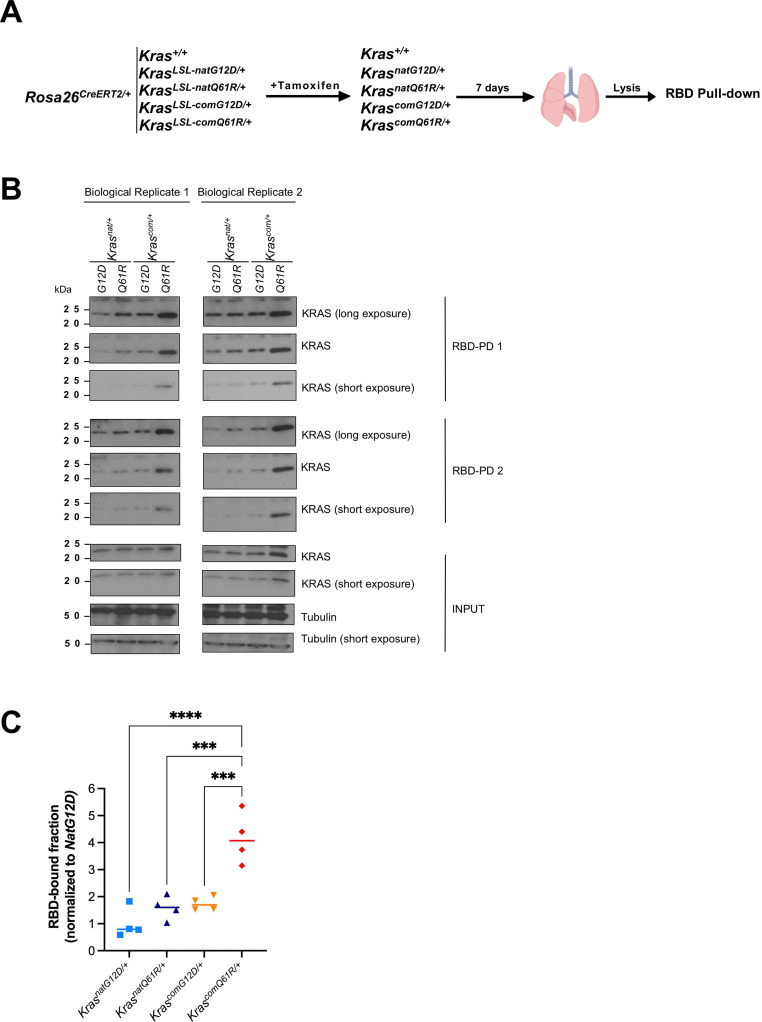
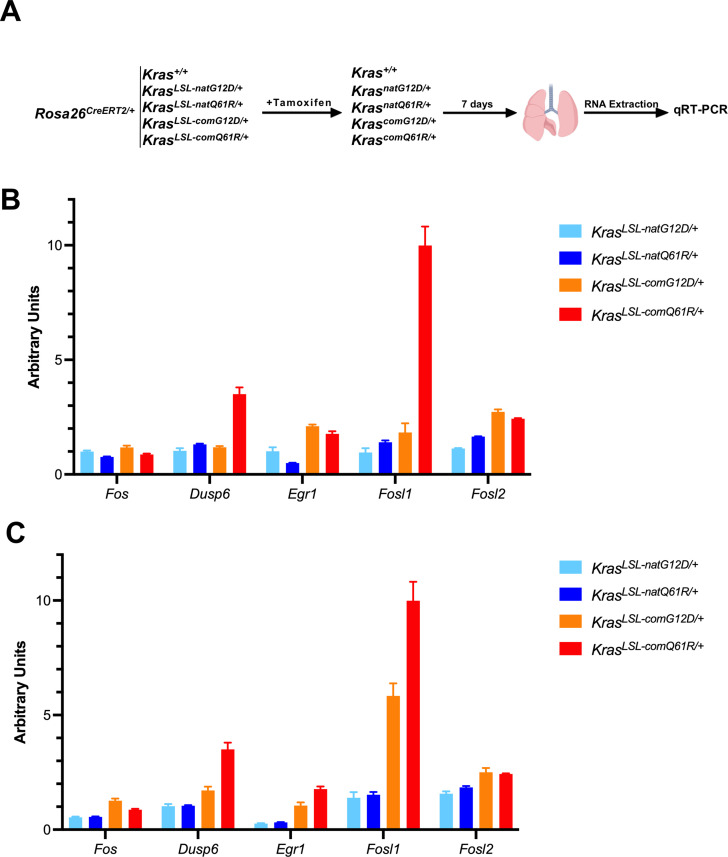
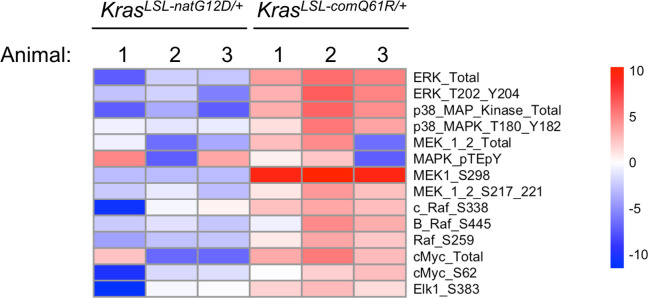
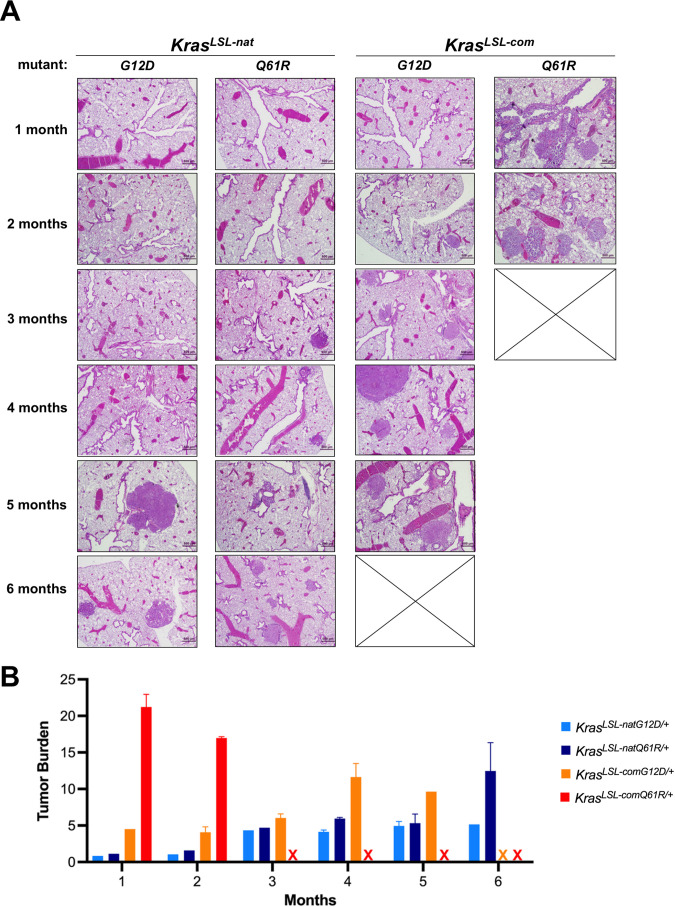
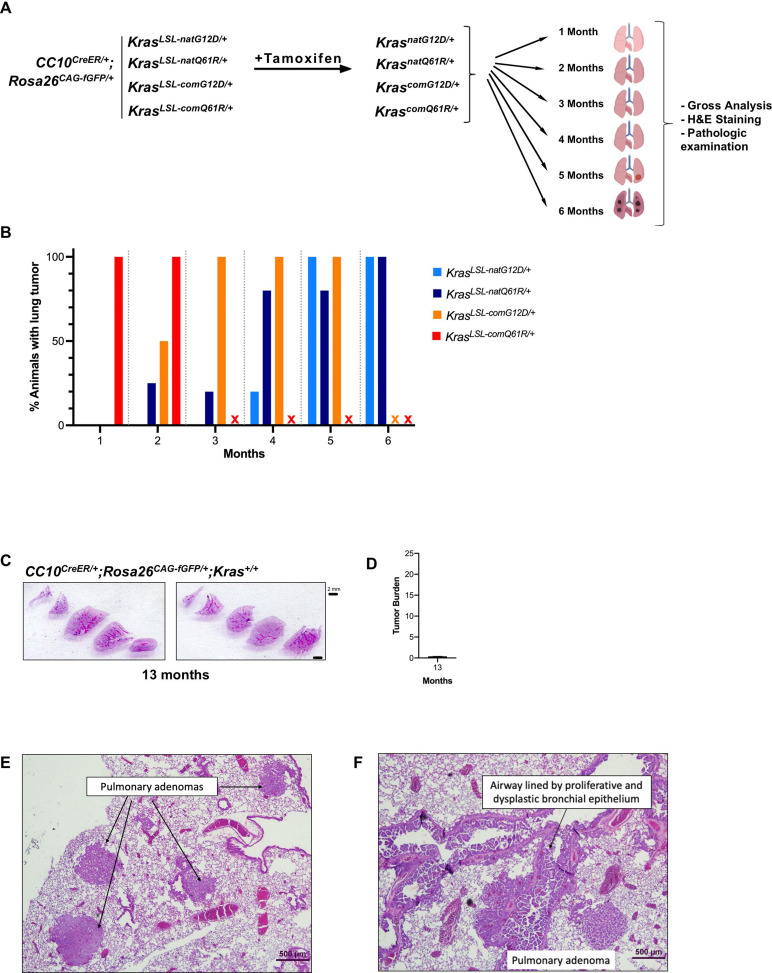
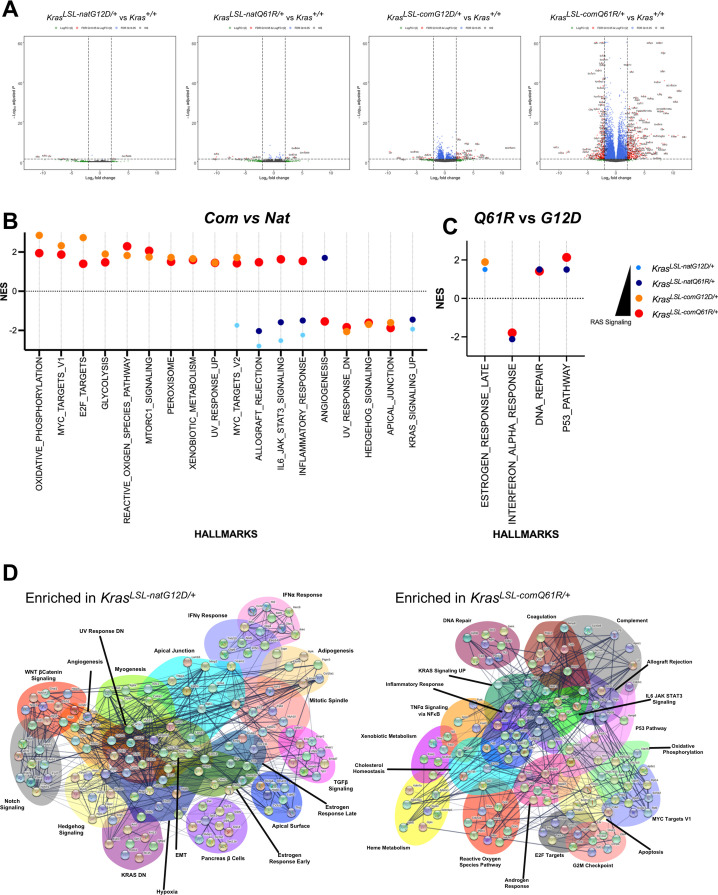
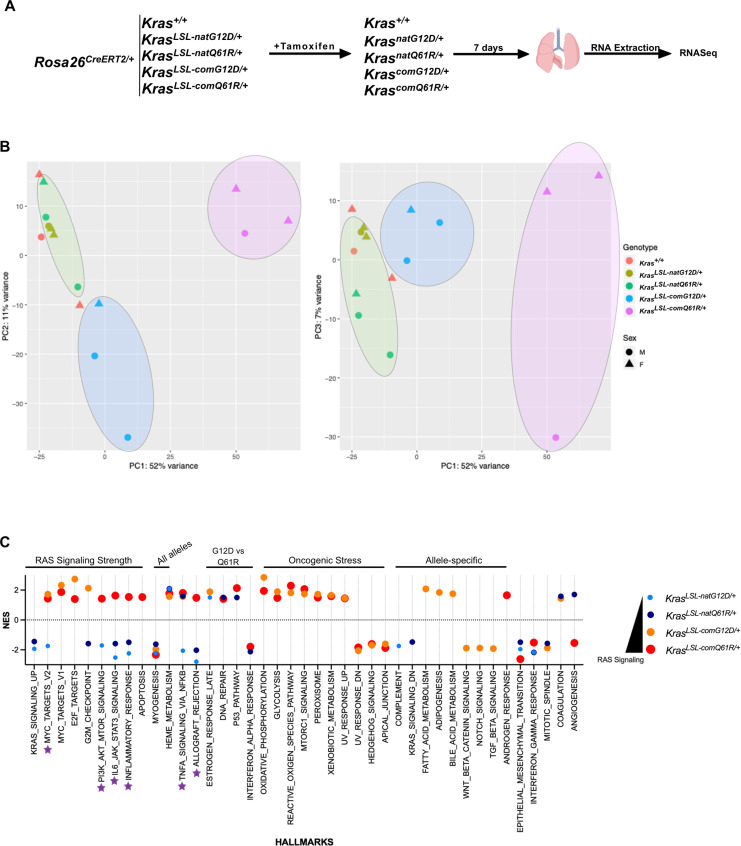
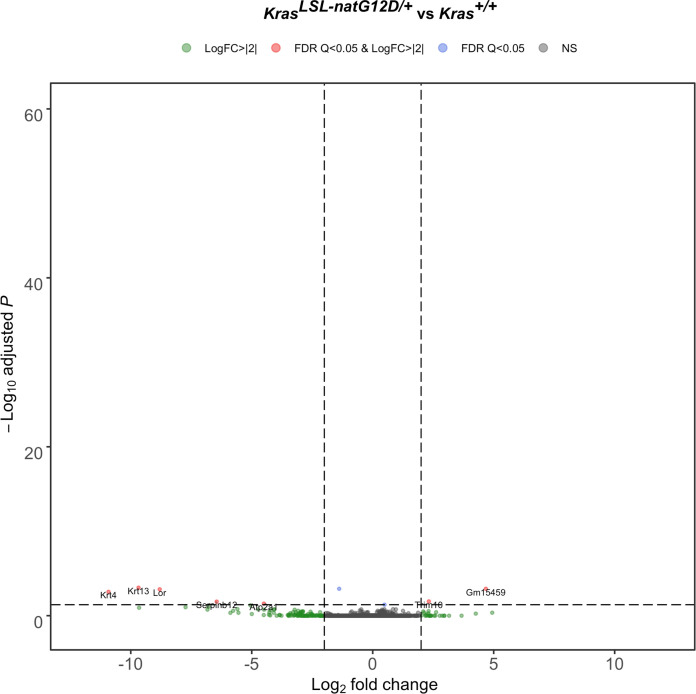
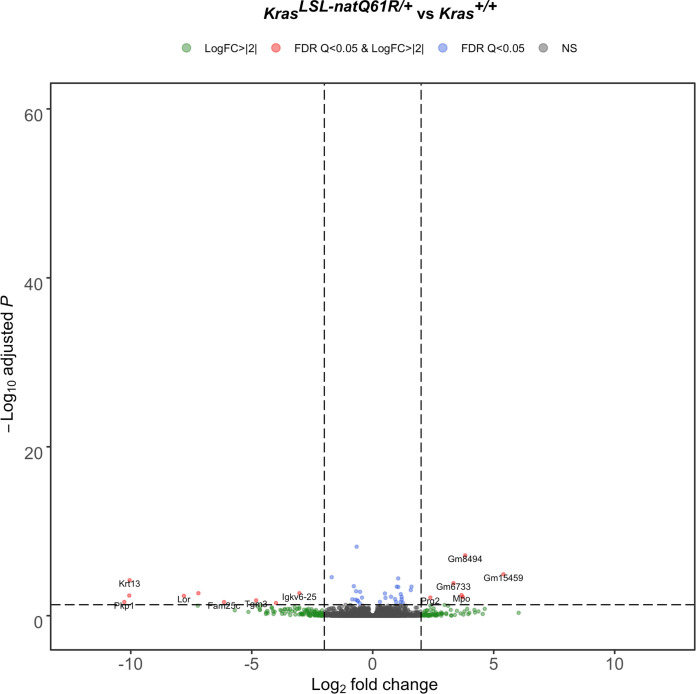
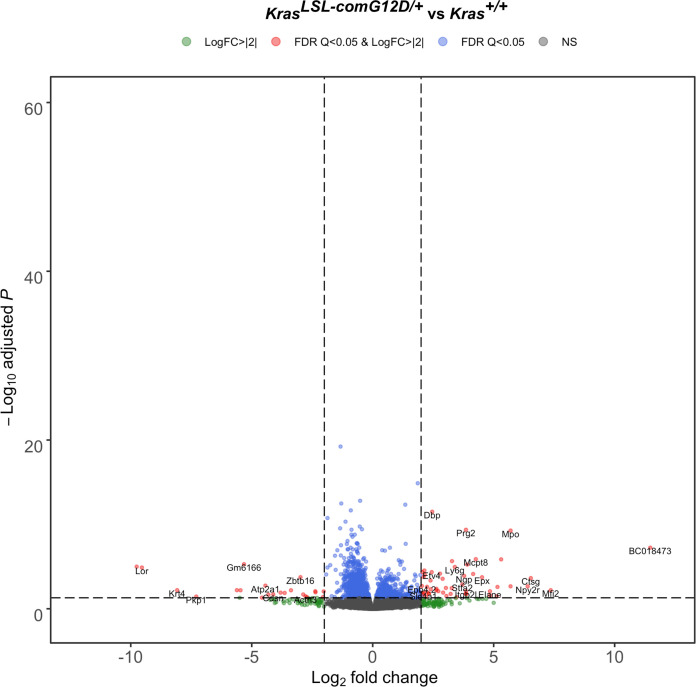
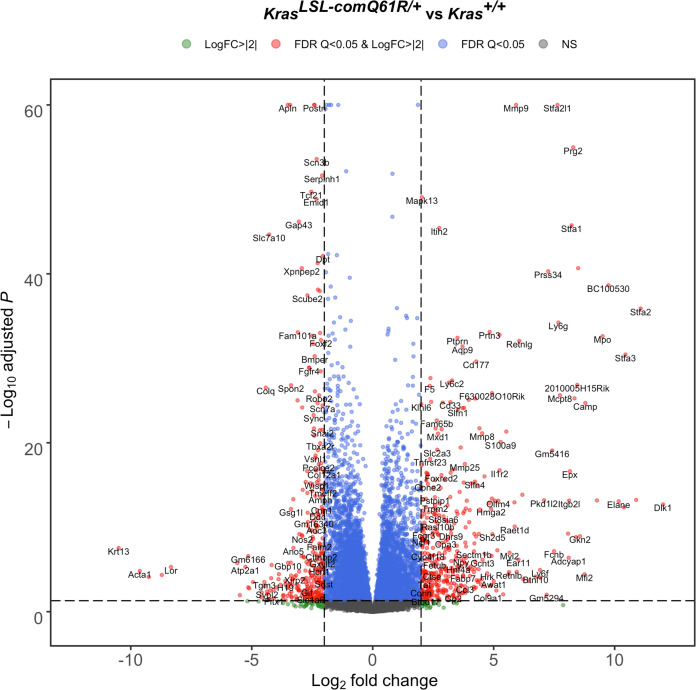
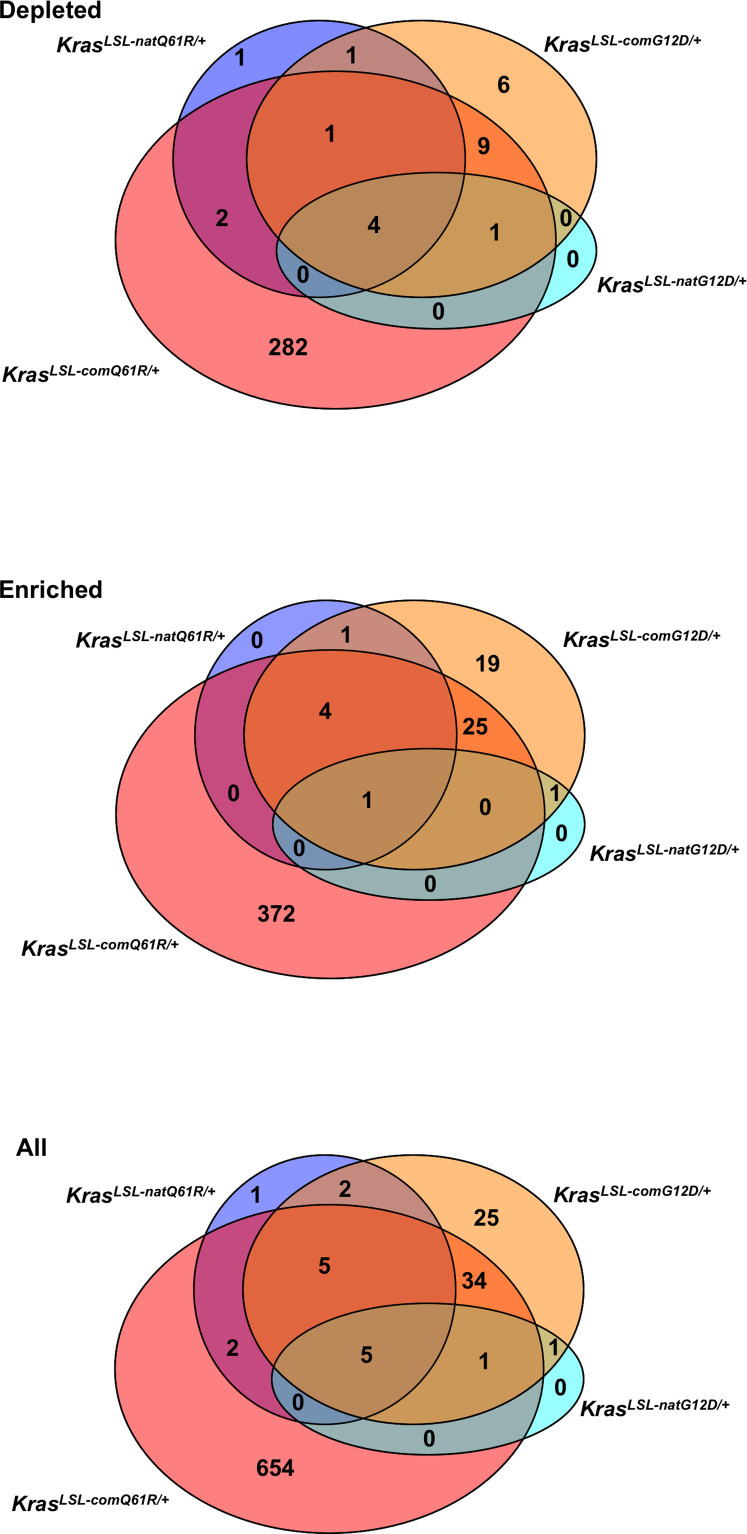
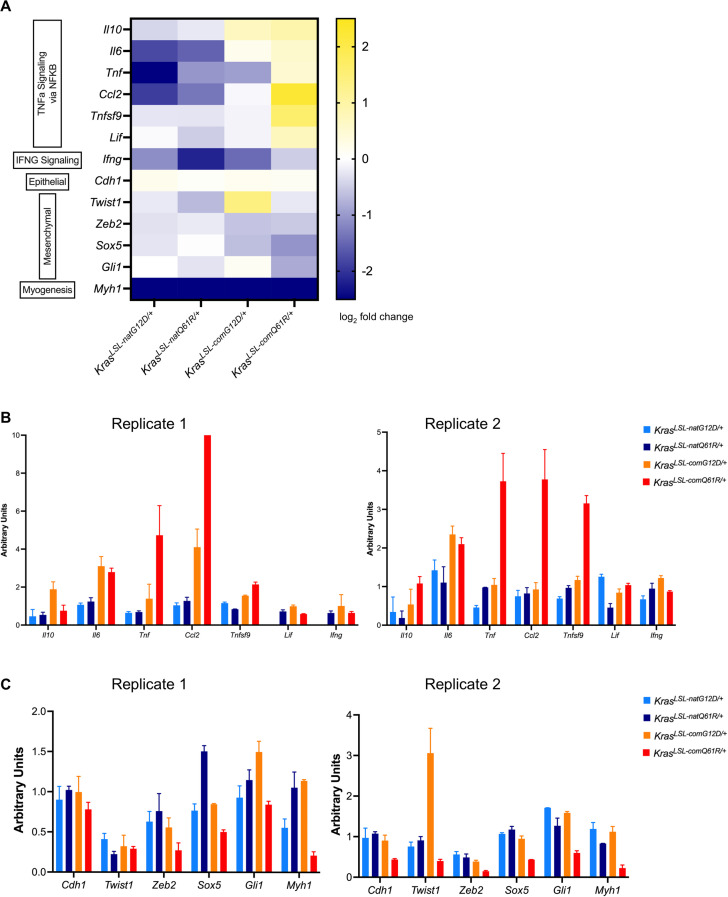
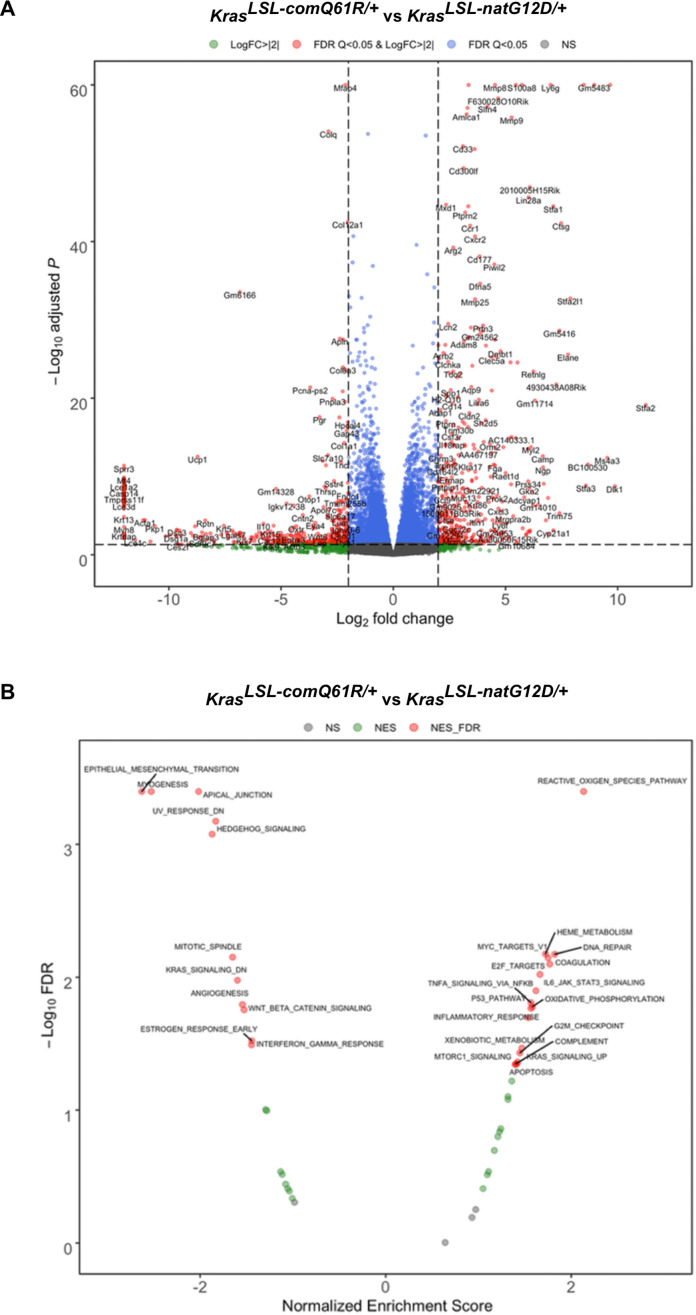
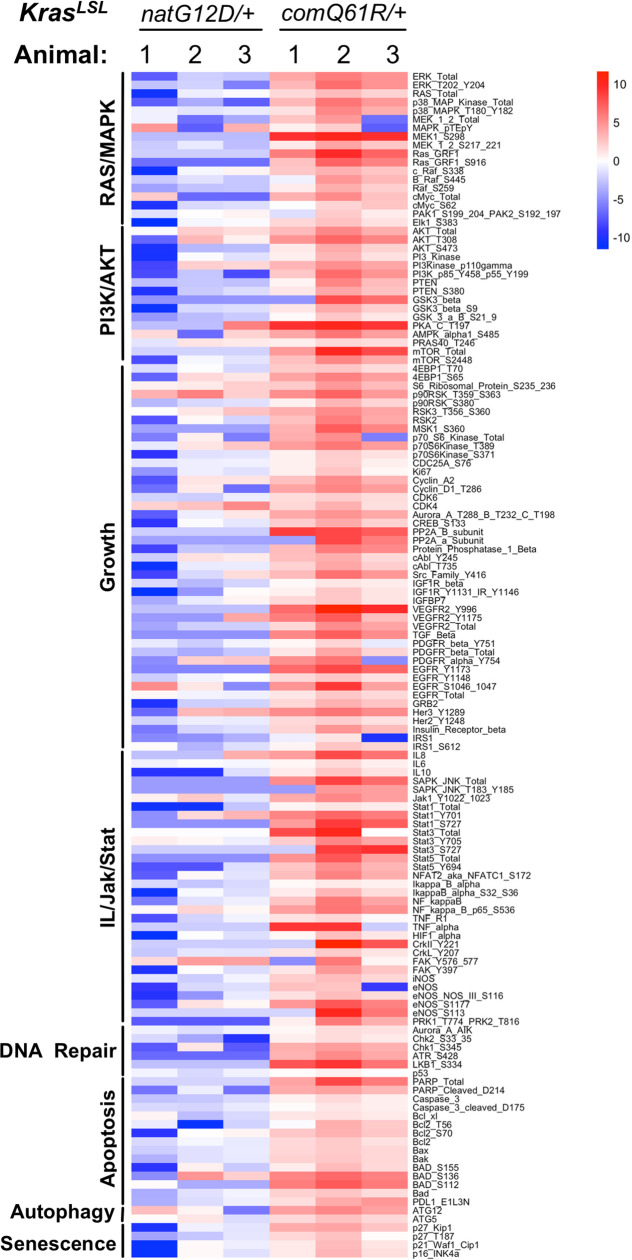
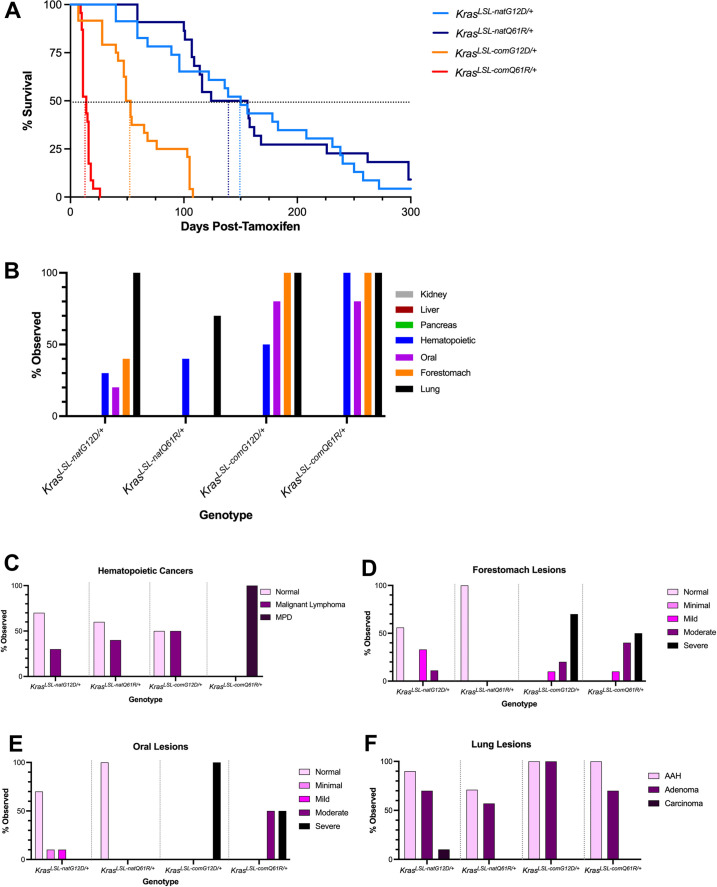
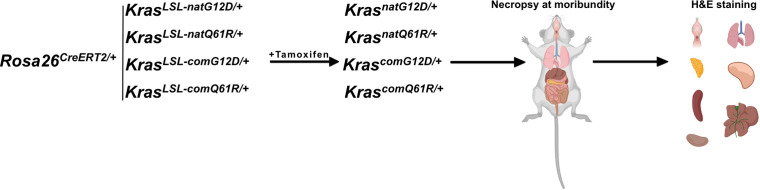
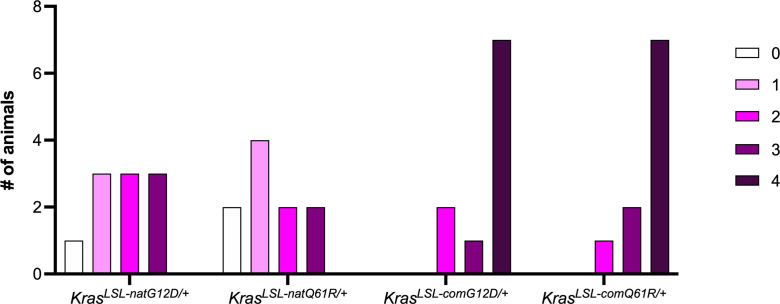
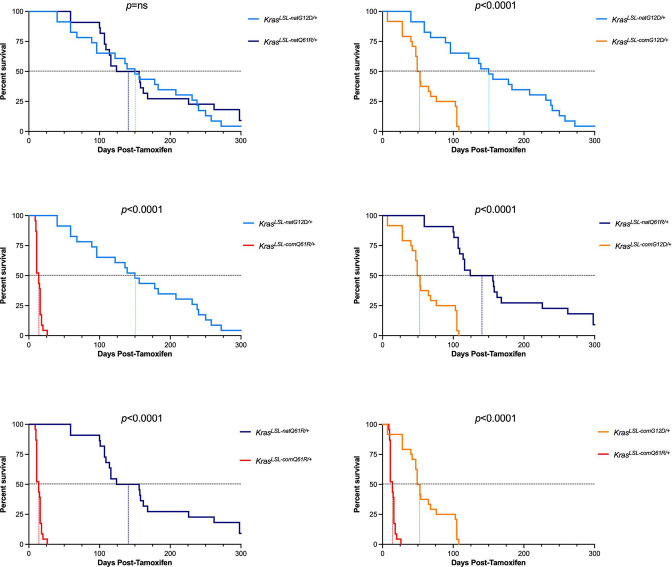
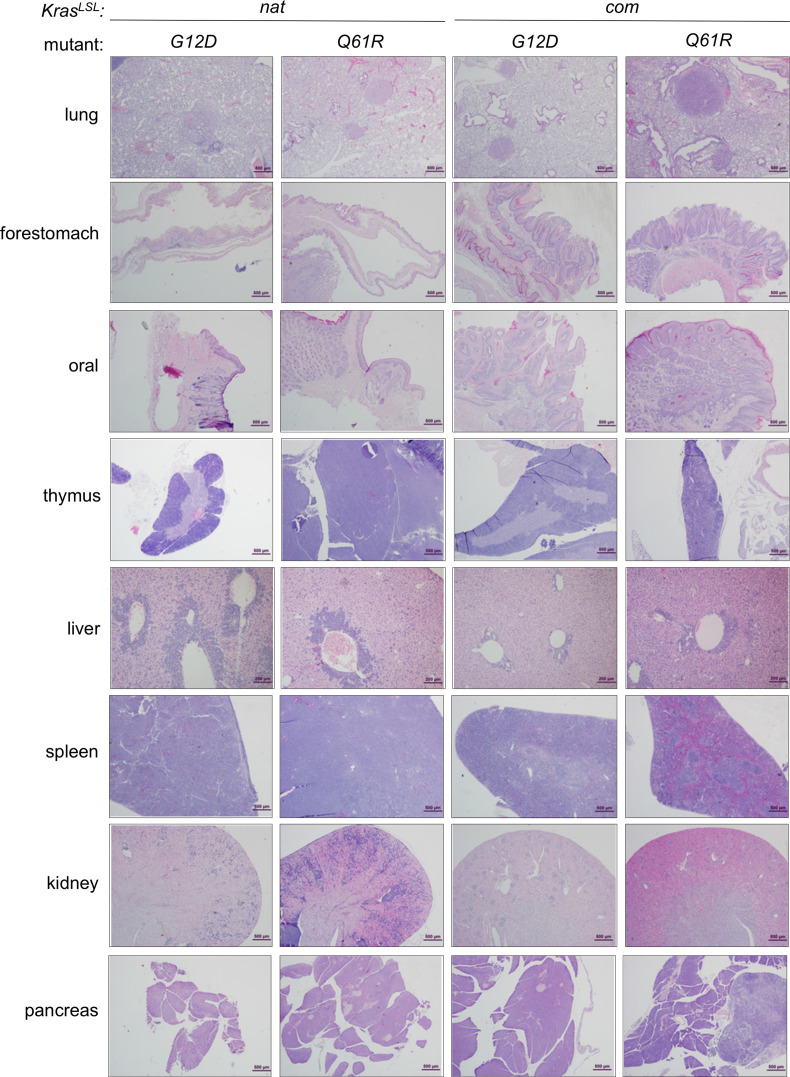
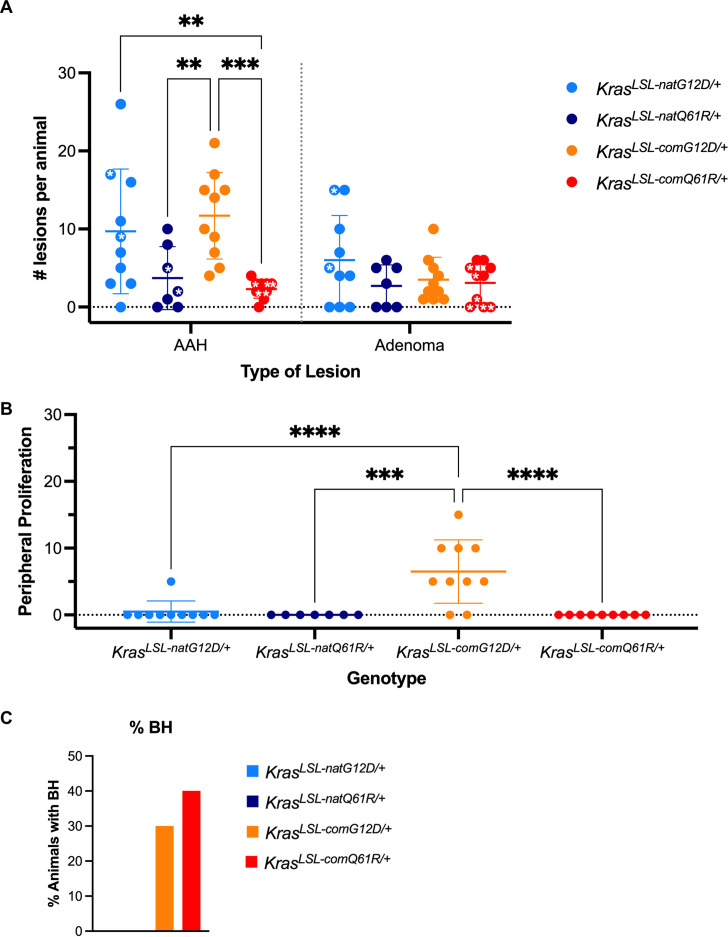
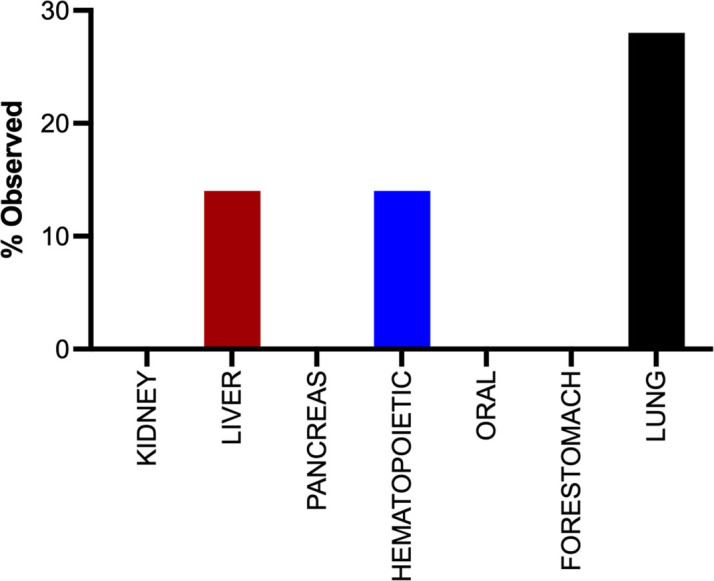
Despite multiple possible oncogenic mutations in the proto-oncogene , unique subsets of these mutations are detected in different cancer types. As mutations occur early, if not being the initiating event, these mutational biases are ostensibly a product of how normal cells respond to the encoded oncoprotein. Oncogenic mutations can impact not only the level of active oncoprotein, but also engagement with proteins. To attempt to separate these two effects, we generated four novel Cre-inducible (LSL) alleles in mice with the biochemically distinct G12D or Q61R mutations and encoded by native (nat) rare or common (com) codons to produce low or high protein levels. While there were similarities, each allele also induced a distinct transcriptional response shortly after activation in vivo. At one end of the spectrum, activating the allele induced transcriptional hallmarks suggestive of an expansion of multipotent cells, while at the other end, activating the allele led to hallmarks of hyperproliferation and oncogenic stress. Evidence suggests that these changes may be a product of signaling differences due to increased protein expression as well as the specific mutation. To determine the impact of these distinct responses on RAS mutational patterning in vivo, all four alleles were globally activated, revealing that hematolymphopoietic lesions were permissive to the level of active oncoprotein, squamous tumors were permissive to the G12D mutant, while carcinomas were permissive to both these features. We suggest that different KRAS mutations impart unique signaling properties that are preferentially capable of inducing tumor initiation in a distinct cell-specific manner.
尽管原癌基因中有多种可能的致癌突变,但在不同的癌症类型中仅检测到这些突变的独特子集。由于 突变发生得很早,如果不是起始事件,这些突变偏向显然是正常细胞对编码致癌蛋白的反应的产物。致癌突变不仅会影响活性致癌蛋白的水平,还会影响与蛋白质的结合。为了尝试将这两种效应分开,我们在小鼠中生成了四个具有不同生化特征的新型 Cre 诱导(LSL) 等位基因,这些等位基因具有独特的 G12D 或 Q61R 突变,由天然(nat)稀有或常见(com)密码子编码,以产生低或高水平的蛋白质。虽然有相似之处,但每个等位基因在体内激活后不久也会诱导出独特的转录反应。在频谱的一端,激活 等位基因诱导了多能细胞扩张的转录特征,而在另一端,激活 等位基因导致了过度增殖和致癌应激的特征。有证据表明,这些变化可能是由于蛋白质表达增加以及特定突变导致的信号差异的产物。为了确定这些不同反应对体内 RAS 突变模式的影响,我们全局激活了所有四个等位基因,结果表明,造血淋巴系统病变对活性致癌蛋白的水平是允许的,鳞状肿瘤对 G12D 突变是允许的,而癌是允许这两个特征的。我们认为,不同的 KRAS 突变赋予了独特的信号特性,这些特性能够以独特的细胞特异性方式优先诱导肿瘤起始。