Department of Biochemistry, University of Zurich, Zurich, Switzerland.
Elife. 2022 Sep 8;11:e80332. doi: 10.7554/eLife.80332.
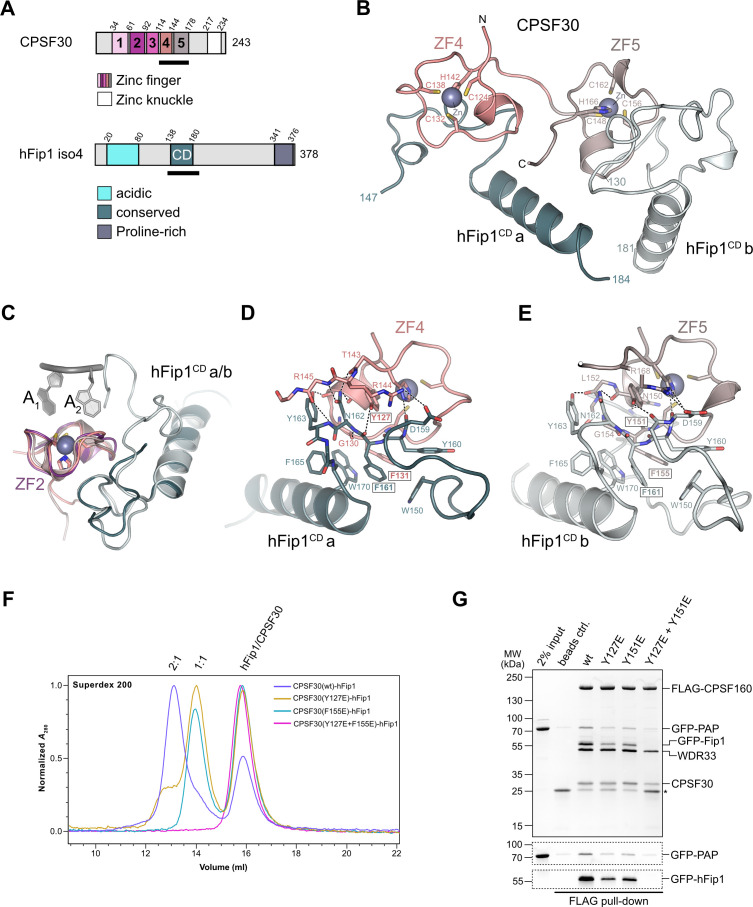
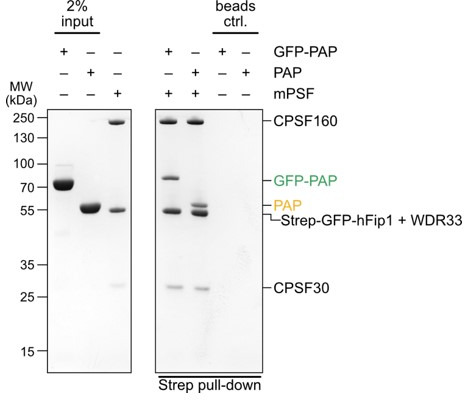
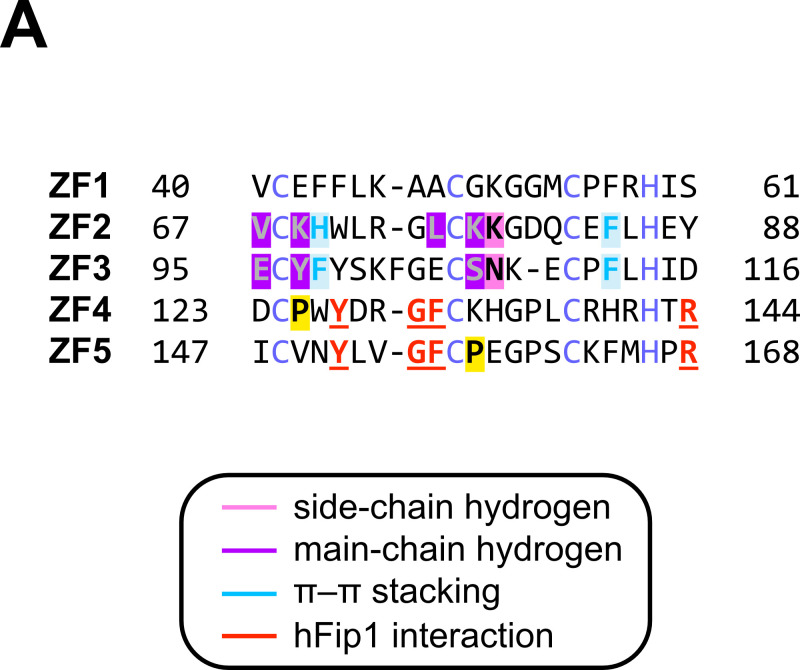
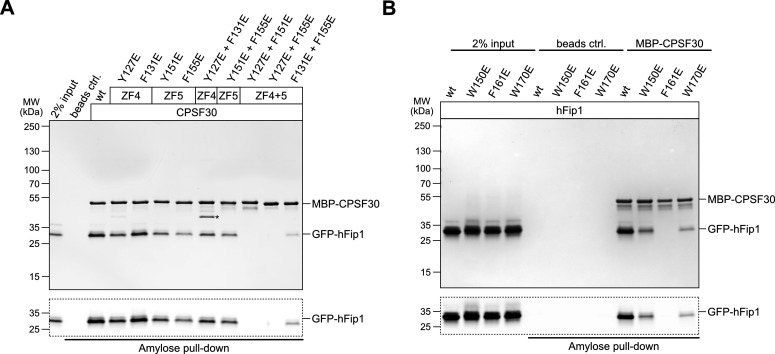
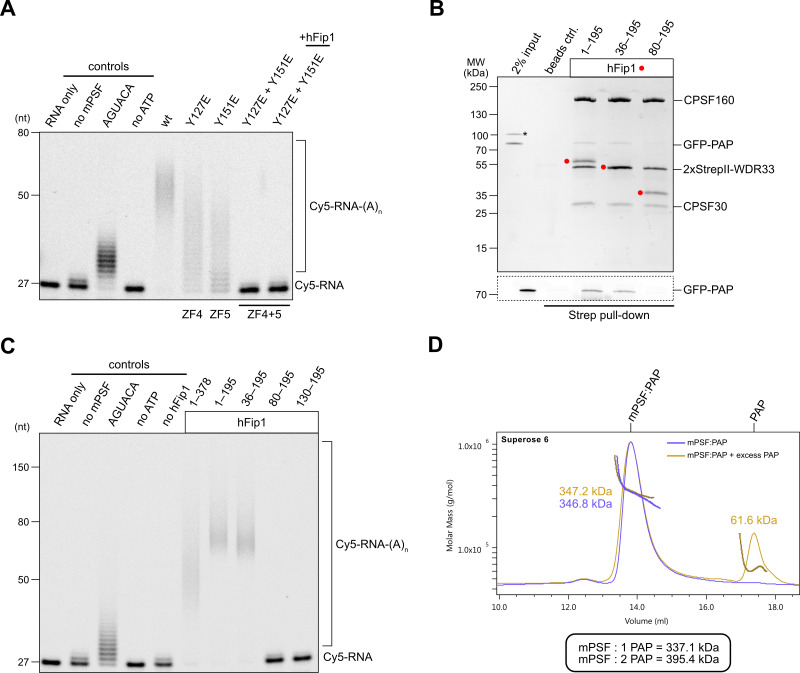
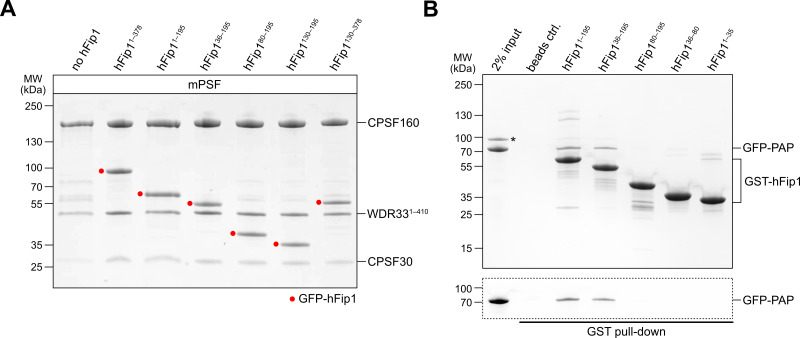
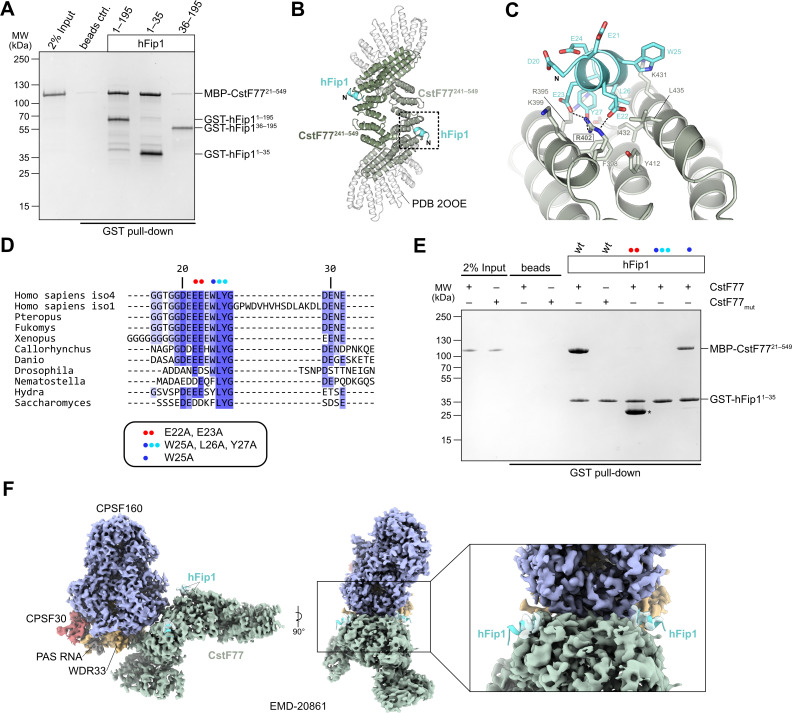
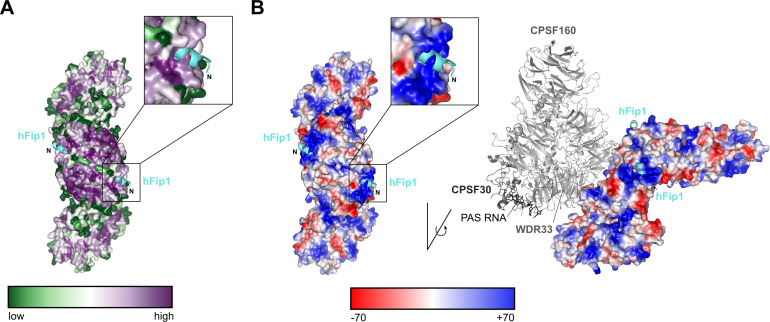
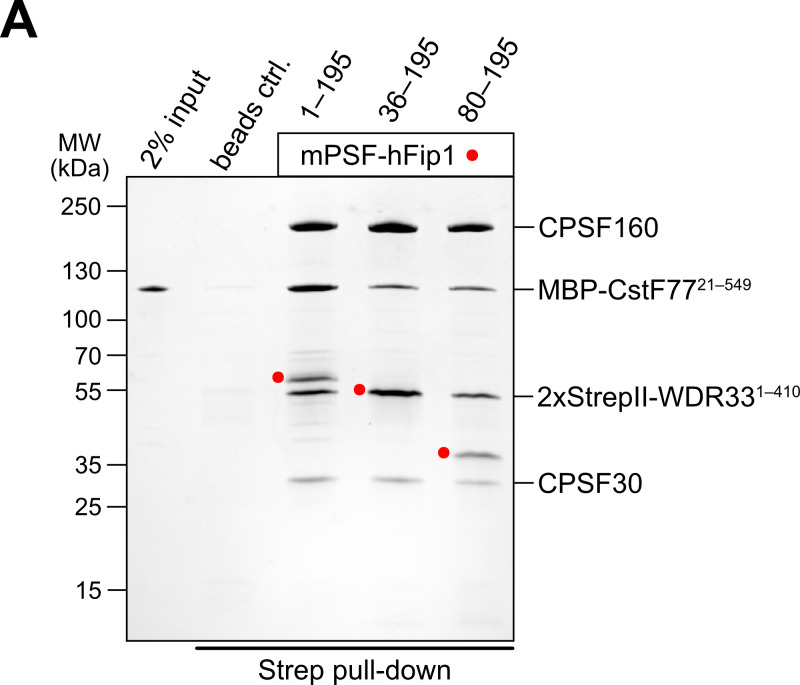
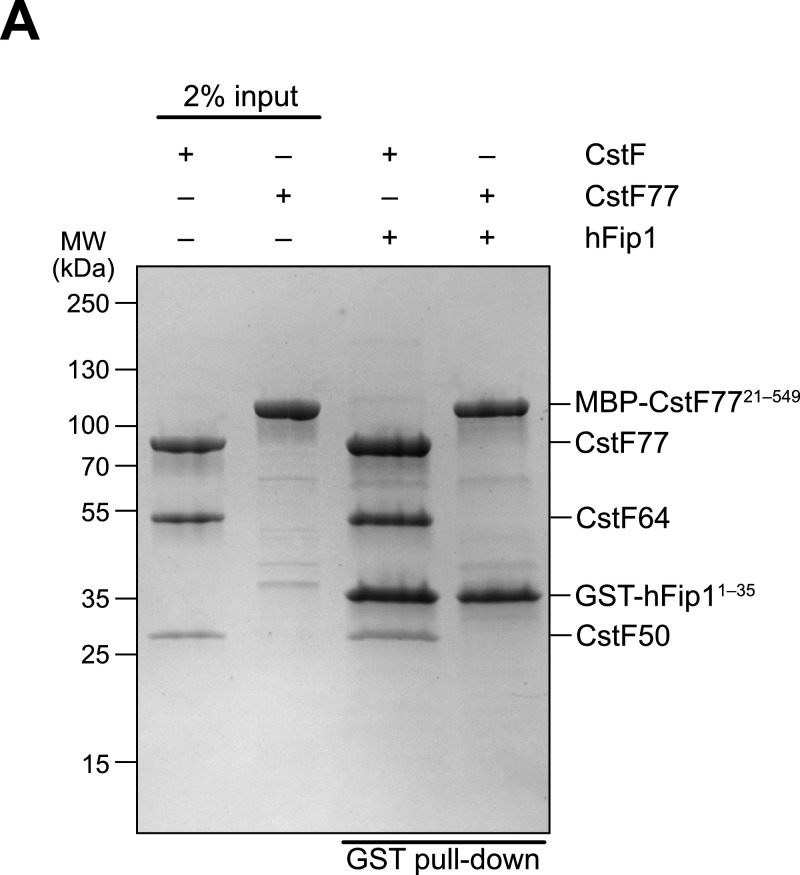
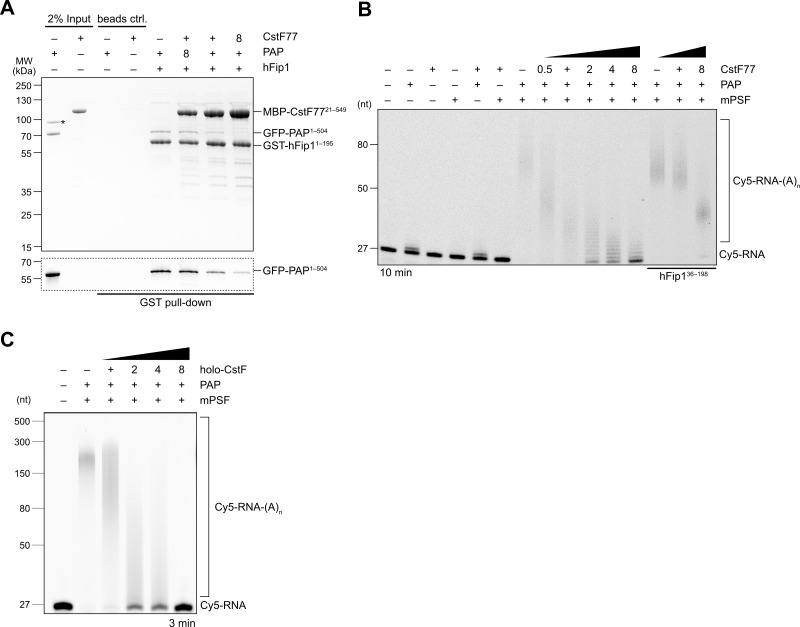
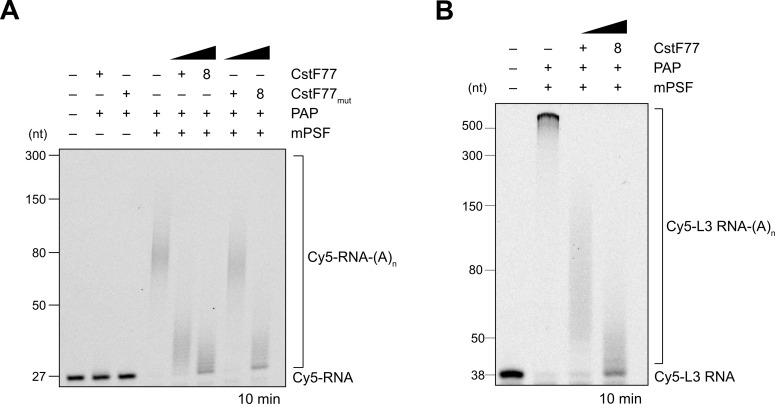
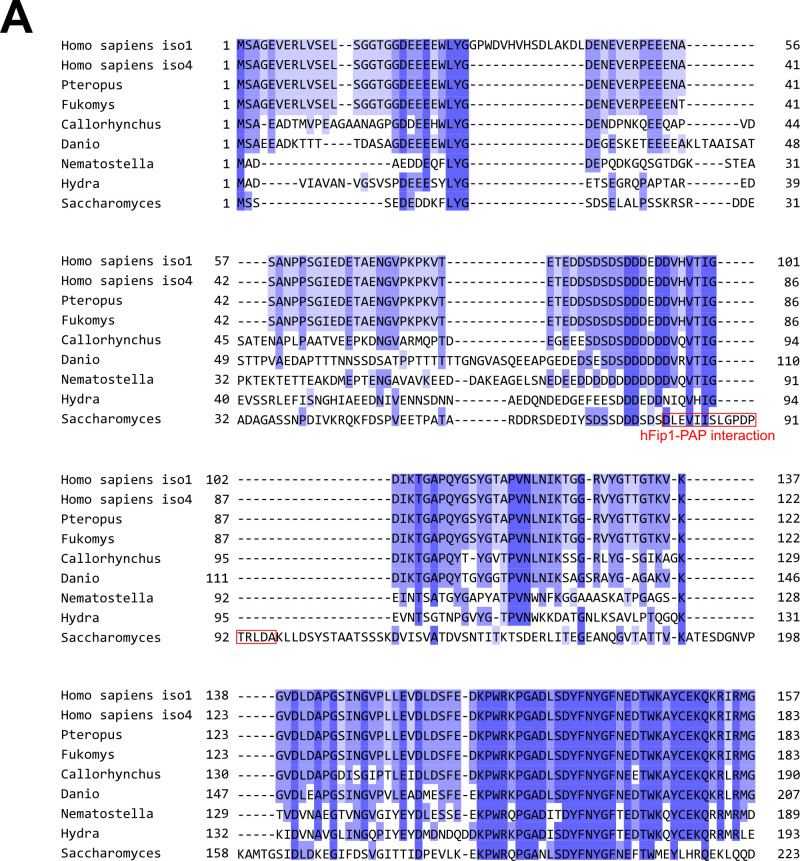
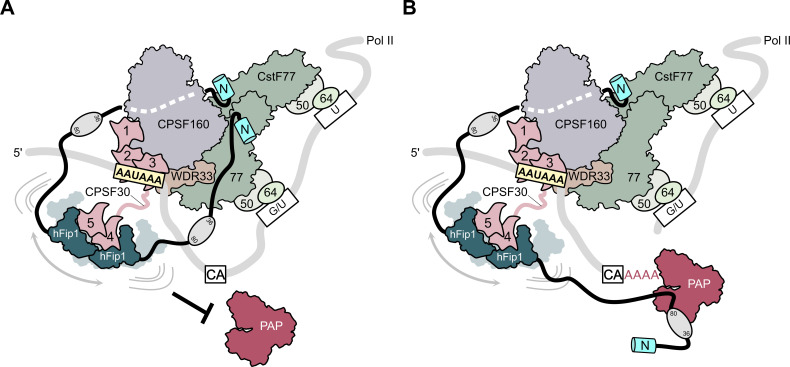
3' end formation of most eukaryotic mRNAs is dependent on the assembly of a ~1.5 MDa multiprotein complex, that catalyzes the coupled reaction of pre-mRNA cleavage and polyadenylation. In mammals, the cleavage and polyadenylation specificity factor (CPSF) constitutes the core of the 3' end processing machinery onto which the remaining factors, including cleavage stimulation factor (CstF) and poly(A) polymerase (PAP), assemble. These interactions are mediated by Fip1, a CPSF subunit characterized by high degree of intrinsic disorder. Here, we report two crystal structures revealing the interactions of human Fip1 (hFip1) with CPSF30 and CstF77. We demonstrate that CPSF contains two copies of hFip1, each binding to the zinc finger (ZF) domains 4 and 5 of CPSF30. Using polyadenylation assays we show that the two hFip1 copies are functionally redundant in recruiting one copy of PAP, thereby increasing the processivity of RNA polyadenylation. We further show that the interaction between hFip1 and CstF77 is mediated via a short motif in the N-terminal 'acidic' region of hFip1. In turn, CstF77 competitively inhibits CPSF-dependent PAP recruitment and 3' polyadenylation. Taken together, these results provide a structural basis for the multivalent scaffolding and regulatory functions of hFip1 in 3' end processing.
3' 端大多数真核 mRNA 的形成依赖于一个约 1.5 MDa 的多蛋白复合物的组装,该复合物催化前体 mRNA 切割和多聚腺苷酸化的偶联反应。在哺乳动物中,切割和多聚腺苷酸化特异性因子(CPSF)构成 3' 端加工机制的核心,其余因子,包括切割刺激因子(CstF)和多聚(A)聚合酶(PAP),组装在这个核心上。这些相互作用由 Fip1 介导,Fip1 是 CPSF 的一个亚基,具有高度的固有无序性。在这里,我们报告了两个晶体结构,揭示了人 Fip1(hFip1)与 CPSF30 和 CstF77 的相互作用。我们证明 CPSF 包含两个 hFip1 拷贝,每个拷贝都与 CPSF30 的锌指(ZF)结构域 4 和 5 结合。我们使用多聚腺苷酸化测定表明,两个 hFip1 拷贝在招募一个 PAP 拷贝方面具有功能冗余性,从而增加了 RNA 多聚腺苷酸化的连续性。我们进一步表明,hFip1 与 CstF77 之间的相互作用是通过 hFip1 的 N 端“酸性”区域中的一个短基序介导的。反过来,CstF77 竞争性地抑制 CPSF 依赖性 PAP 招募和 3' 多聚腺苷酸化。总之,这些结果为 hFip1 在 3' 端加工中的多价支架和调节功能提供了结构基础。