Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan, China.
Elife. 2022 Sep 29;11:e79926. doi: 10.7554/eLife.79926.
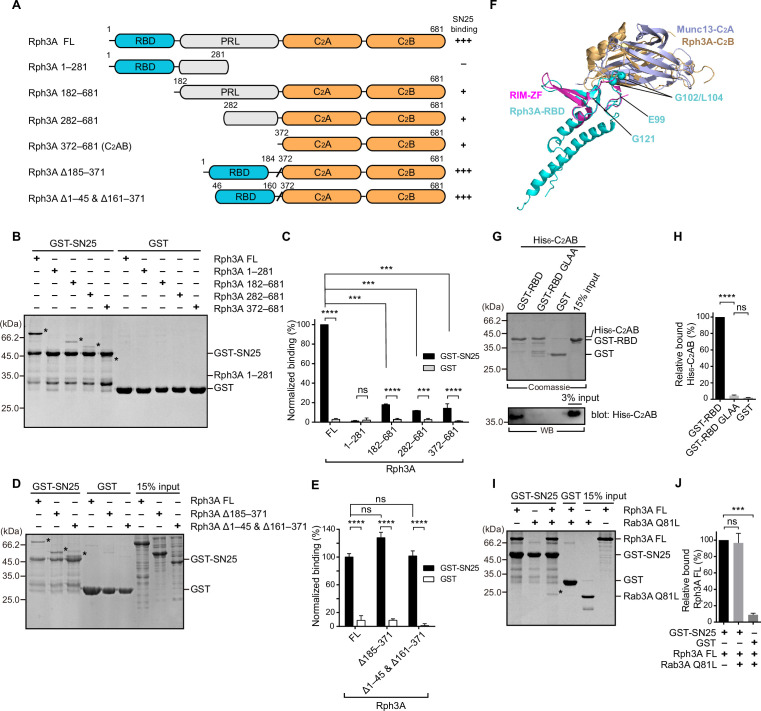
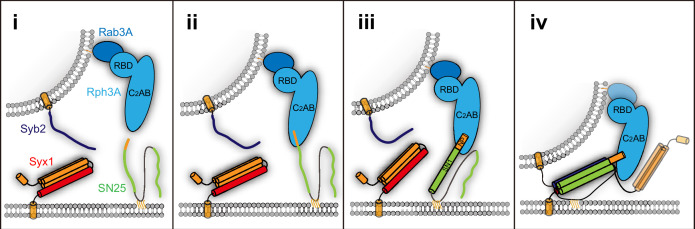
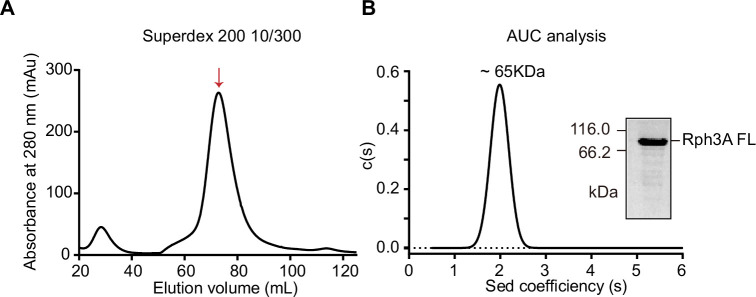
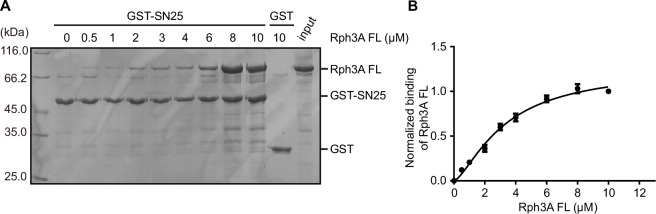
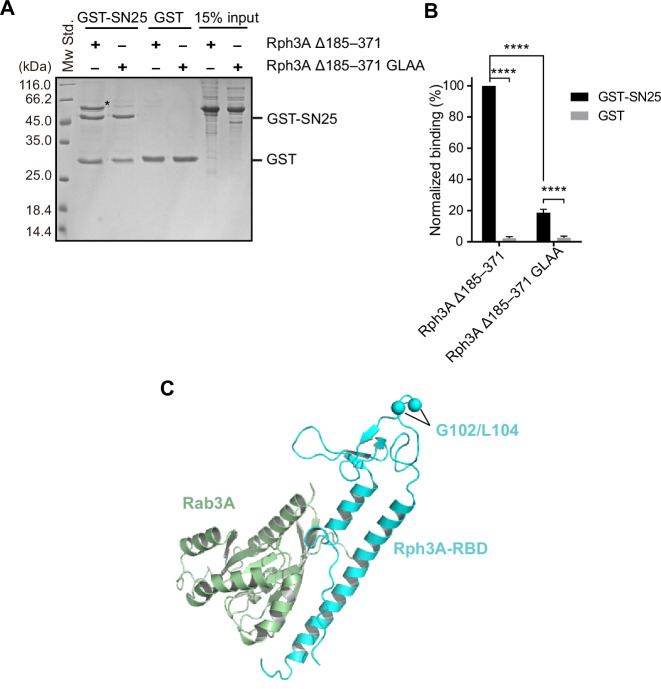
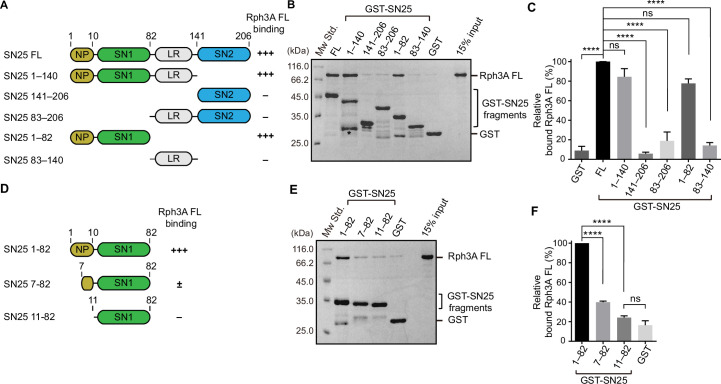
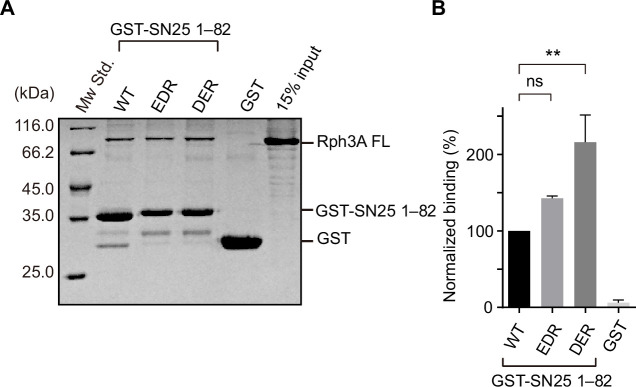
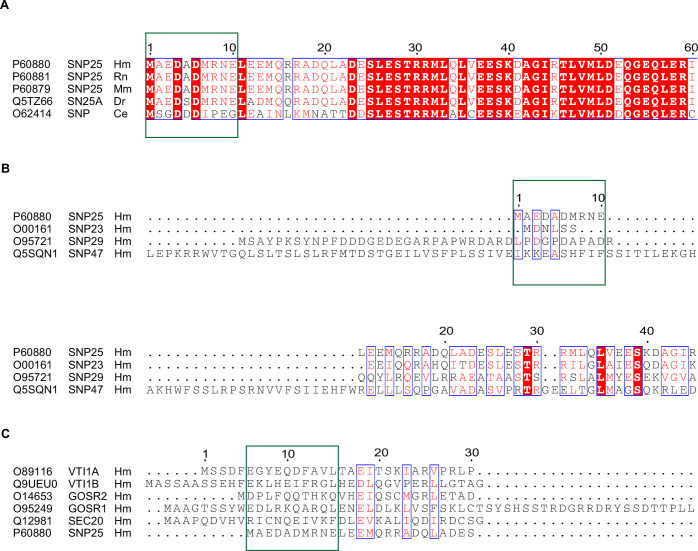
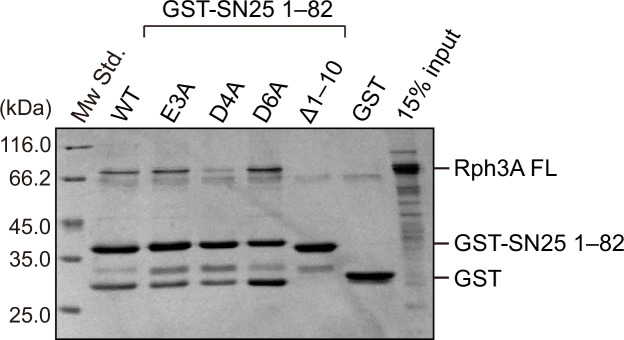
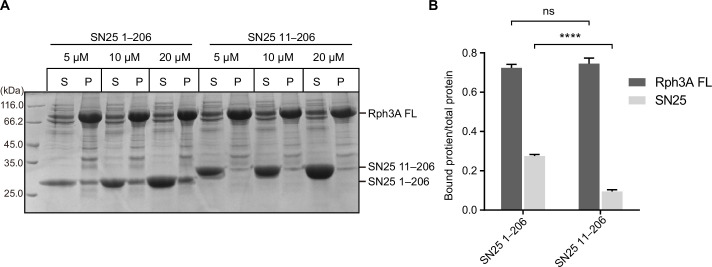
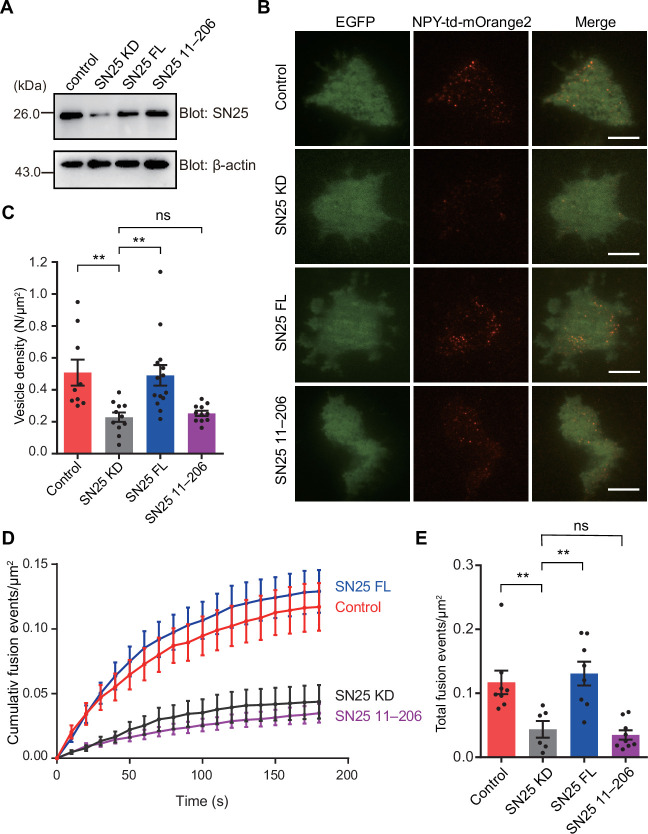
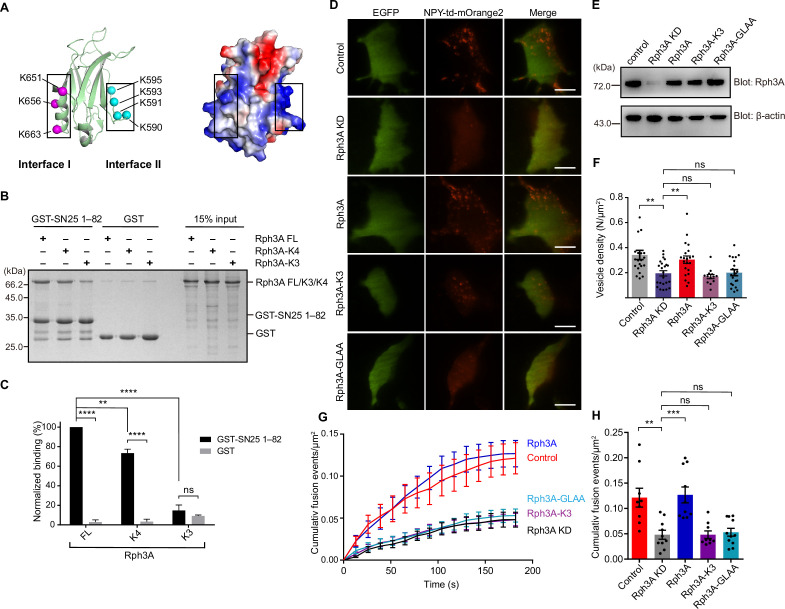
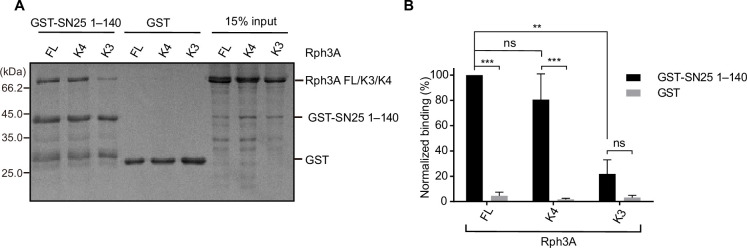
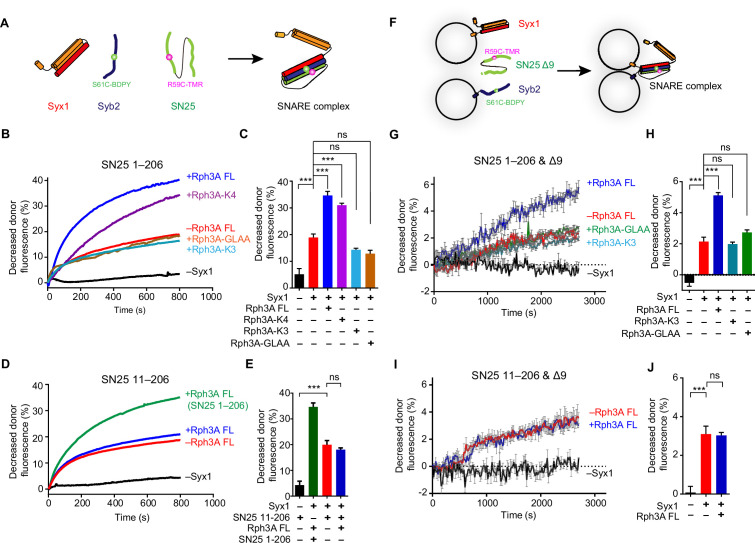
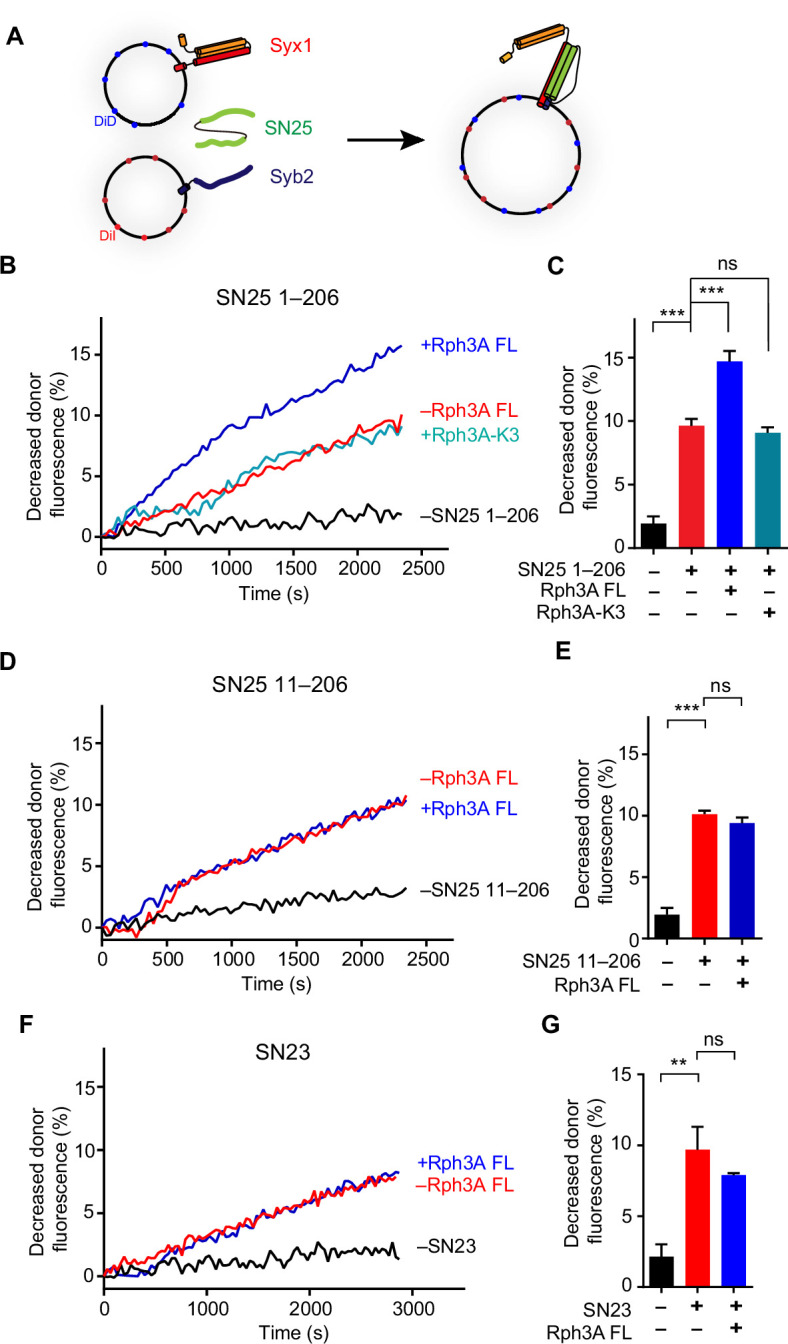
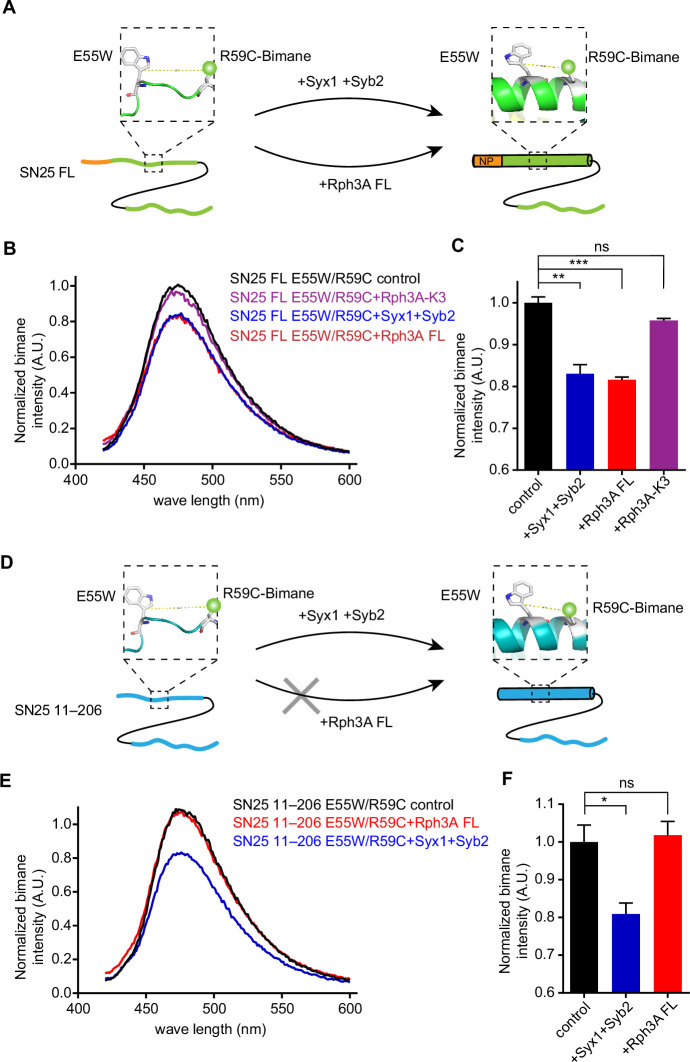
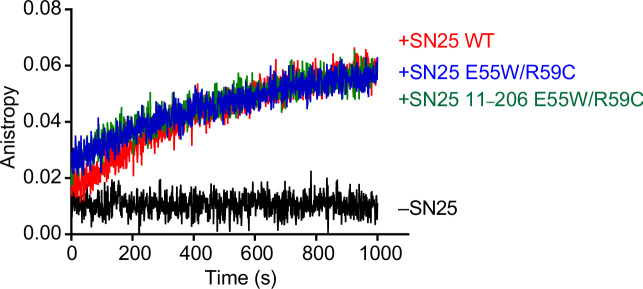
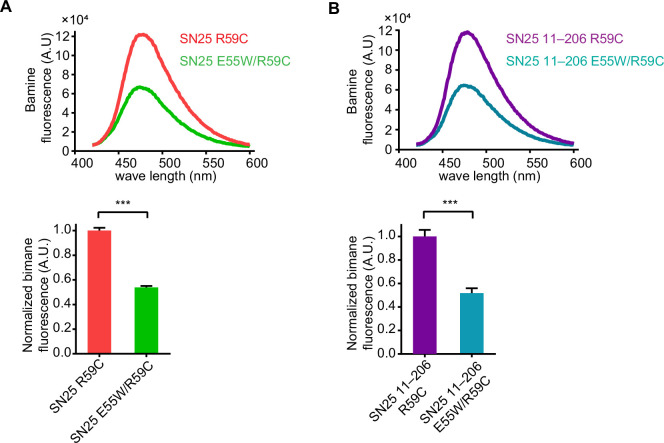
Exocytosis of secretory vesicles requires the soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) proteins and small GTPase Rabs. As a Rab3/Rab27 effector protein on secretory vesicles, Rabphilin 3A was implicated to interact with SNAP-25 to regulate vesicle exocytosis in neurons and neuroendocrine cells, yet the underlying mechanism remains unclear. In this study, we have characterized the physiologically relevant binding sites between Rabphilin 3A and SNAP-25. We found that an intramolecular interplay between the N-terminal Rab-binding domain and C-terminal CAB domain enables Rabphilin 3A to strongly bind the SNAP-25 N-peptide region via its CB bottom α-helix. Disruption of this interaction significantly impaired docking and fusion of vesicles with the plasma membrane in rat PC12 cells. In addition, we found that this interaction allows Rabphilin 3A to accelerate SNARE complex assembly. Furthermore, we revealed that this interaction accelerates SNARE complex assembly via inducing a conformational switch from random coils to α-helical structure in the SNAP-25 SNARE motif. Altogether, our data suggest that the promotion of SNARE complex assembly by binding the CB bottom α-helix of Rabphilin 3A to the N-peptide of SNAP-25 underlies a pre-fusion function of Rabphilin 3A in vesicle exocytosis.
囊泡的胞吐作用需要可溶性 N-乙基马来酰亚胺敏感因子附着蛋白受体(SNARE)蛋白和小 GTPase Rab 。Rabphilin 3A 作为囊泡上的 Rab3/Rab27 效应蛋白,被认为与 SNAP-25 相互作用,以调节神经元和神经内分泌细胞中的囊泡胞吐作用,但潜在的机制尚不清楚。在本研究中,我们对 Rabphilin 3A 和 SNAP-25 之间的生理相关结合位点进行了表征。我们发现,Rabphilin 3A 中 N 端 Rab 结合域和 C 端 CAB 域之间的分子内相互作用,使其通过其 CB 底部α-螺旋,强烈结合 SNAP-25 N 肽区。破坏这种相互作用会显著损害 PC12 细胞中囊泡与质膜的对接和融合。此外,我们发现这种相互作用允许 Rabphilin 3A 加速 SNARE 复合物的组装。此外,我们发现这种相互作用通过诱导 SNAP-25 SNARE 基序中的随机卷曲到α-螺旋结构的构象转变,来加速 SNARE 复合物的组装。总之,我们的数据表明,Rabphilin 3A 通过结合其 CB 底部α-螺旋与 SNAP-25 的 N 肽,促进 SNARE 复合物的组装,这是 Rabphilin 3A 在囊泡胞吐作用中的一个融合前功能。