Key Laboratory of Cell Proliferation and Differentiation of the Ministry of Education, School of Life Sciences, Peking-Tsinghua Center for Life Sciences, Peking University, Beijing, 100871, China.
Westlake Laboratory of Life Sciences and Biomedicine, Key Laboratory of Structural Biology of Zhejiang Province, School of Life Sciences, Westlake University, 18 Shilongshan Road, Hangzhou, 310024, Zhejiang Province, China.
Genome Biol. 2022 Nov 28;23(1):246. doi: 10.1186/s13059-022-02812-w.
Mammalian cells have three types of RNA polymerases (Pols), Pol I, II, and III. However, the extent to which these polymerases are cross-regulated and the underlying mechanisms remain unclear.
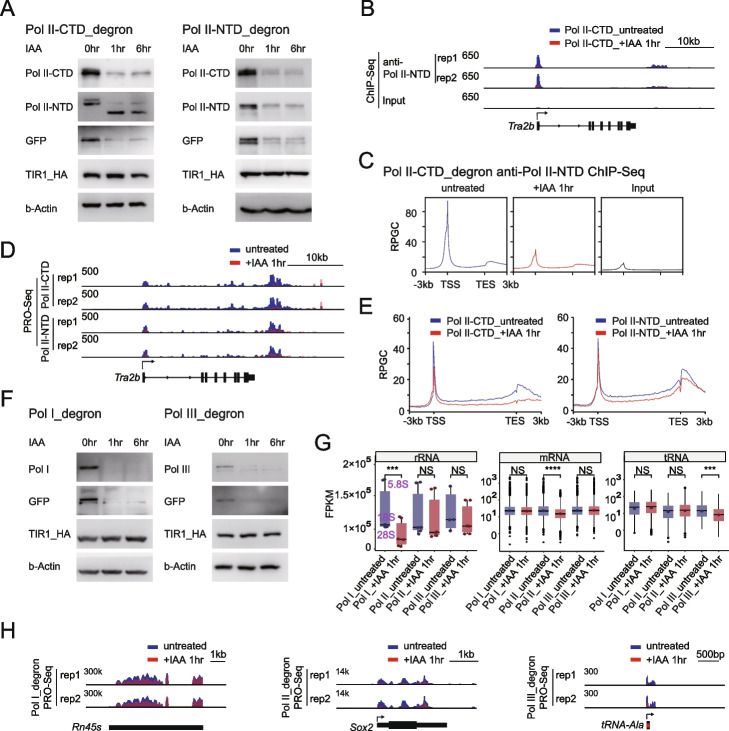
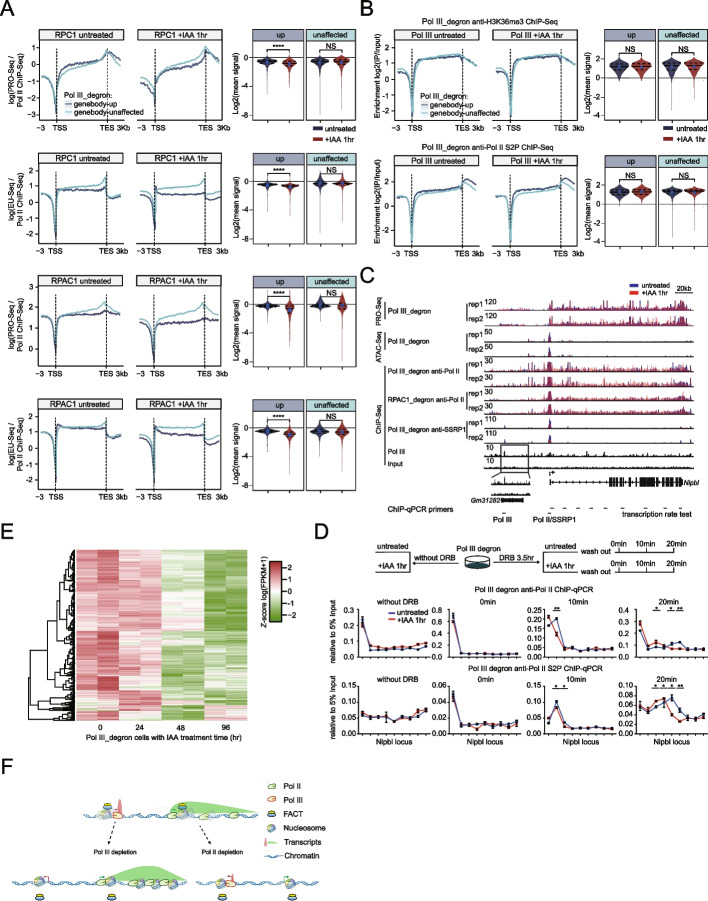
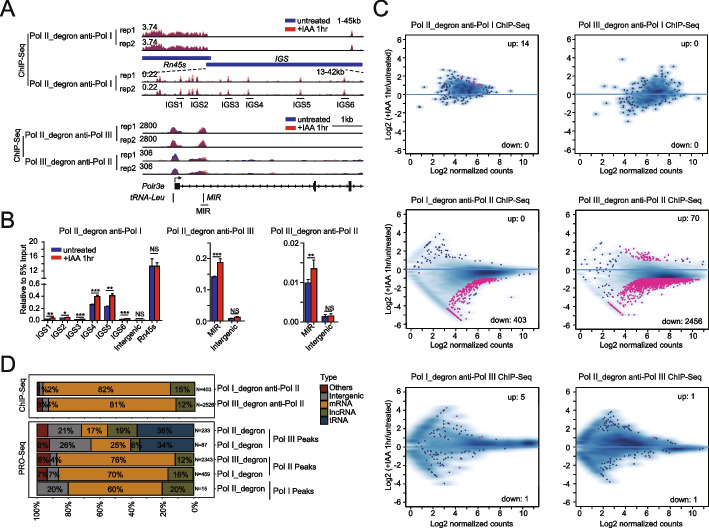
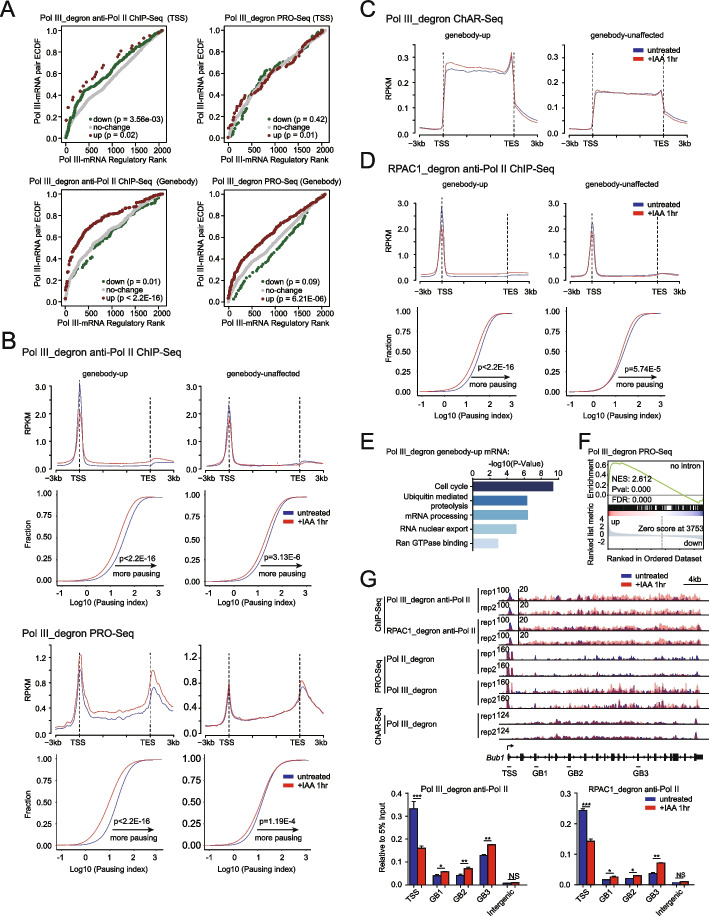
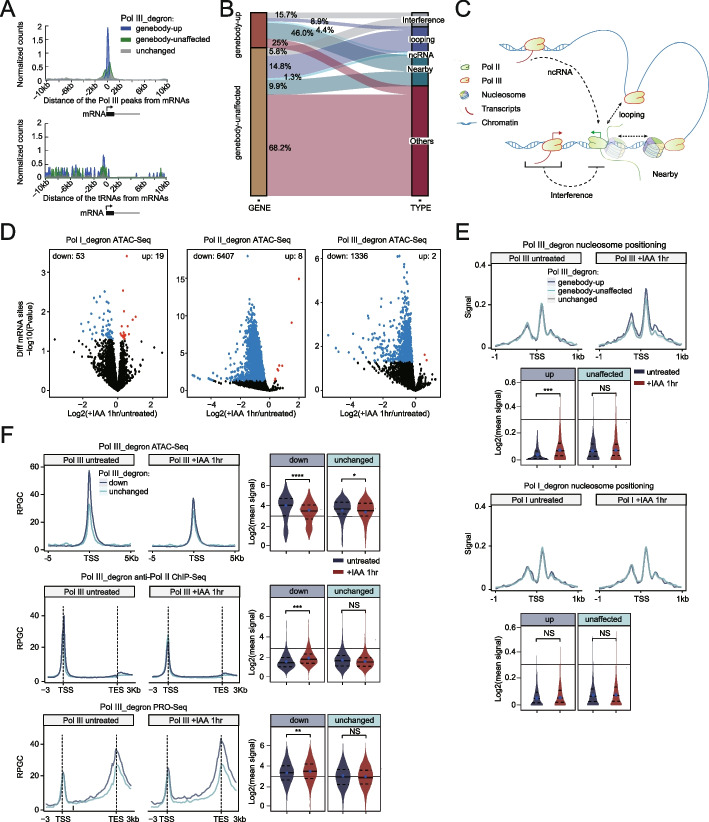
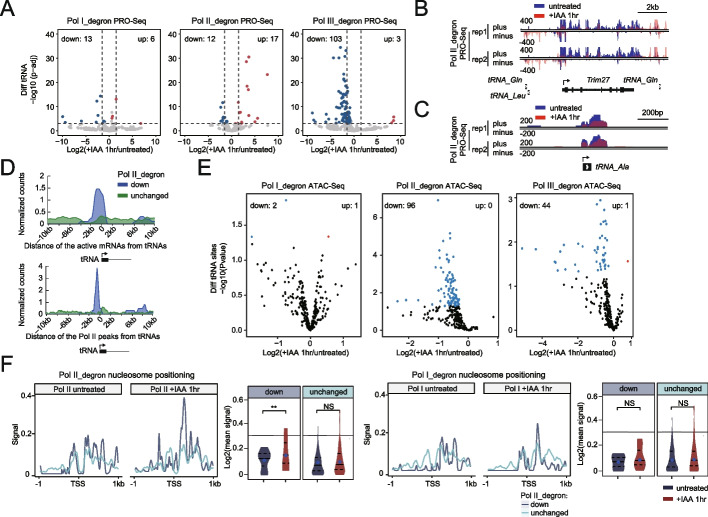
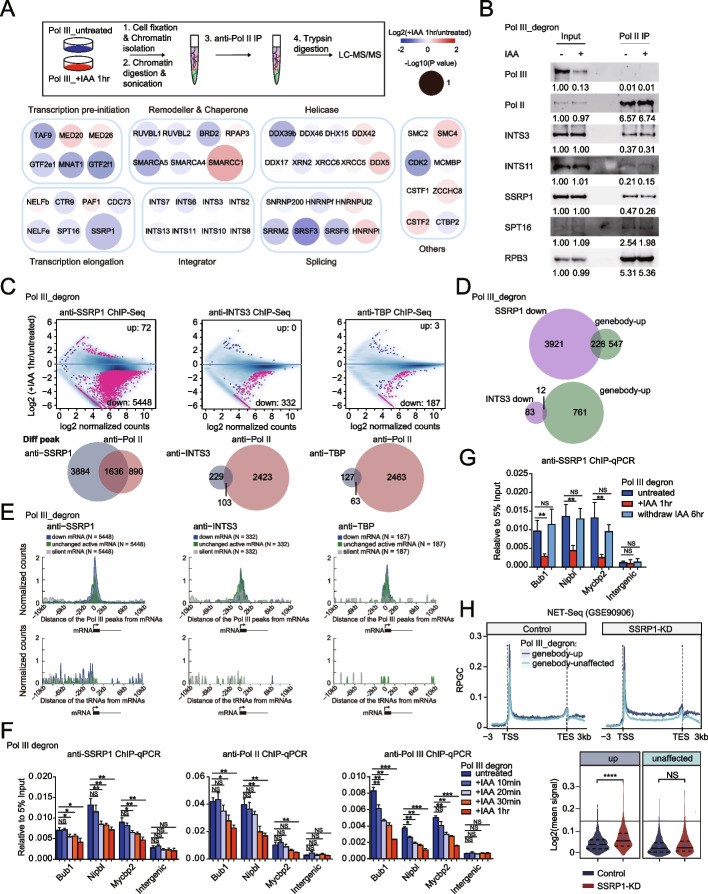
We employ genome-wide profiling after acute depletion of Pol I, Pol II, or Pol III to assess cross-regulatory effects between these Pols. We find that these enzymes mainly affect the transcription of their own target genes, while certain genes are transcribed by the other polymerases. Importantly, the most active type of crosstalk is exemplified by the fact that Pol III depletion affects Pol II transcription. Pol II genes with transcription changes upon Pol III depletion are enriched in diverse cellular functions, and Pol III binding sites are found near their promoters. However, these Pol III binding sites do not correspond to transfer RNAs. Moreover, we demonstrate that Pol III regulates Pol II transcription and chromatin binding of the facilitates chromatin transcription (FACT) complex to alter local chromatin structures, which in turn affects the Pol II transcription rate.
Our results support a model suggesting that RNA polymerases show cross-regulatory effects: Pol III affects local chromatin structures and the FACT-Pol II axis to regulate the Pol II transcription rate at certain gene loci. This study provides a new perspective for understanding the dysregulation of Pol III in various tissues affected by developmental diseases.
哺乳动物细胞有三种 RNA 聚合酶(Pol),即 Pol I、II 和 III。然而,这些聚合酶之间的交叉调控程度及其潜在机制尚不清楚。
我们采用急性耗尽 Pol I、Pol II 或 Pol III 后进行全基因组分析,以评估这些聚合酶之间的交叉调控效应。我们发现这些酶主要影响其自身靶基因的转录,而某些基因则由其他聚合酶转录。重要的是,最活跃的串扰类型是 Pol III 耗尽会影响 Pol II 转录这一事实。在 Pol III 耗尽时转录发生变化的 Pol II 基因在多种细胞功能中富集,并且 Pol III 结合位点位于其启动子附近。然而,这些 Pol III 结合位点并不对应于转移 RNA。此外,我们证明 Pol III 调节 Pol II 转录和促进染色质转录的复合物(FACT)的染色质结合,以改变局部染色质结构,从而影响 Pol II 转录速率。
我们的结果支持这样一种模型,即 RNA 聚合酶表现出交叉调控效应:Pol III 影响局部染色质结构和 FACT-Pol II 轴,以调节某些基因座的 Pol II 转录速率。这项研究为理解发育疾病影响的各种组织中 Pol III 的失调提供了新的视角。