Department of Microbiology, The Ohio State University, Columbus, Ohio, USA.
Department of Chemistry and Chemical Biology, Harvard University, Cambridge, Massachusetts, USA.
mBio. 2023 Feb 28;14(1):e0008923. doi: 10.1128/mbio.00089-23. Epub 2023 Feb 8.
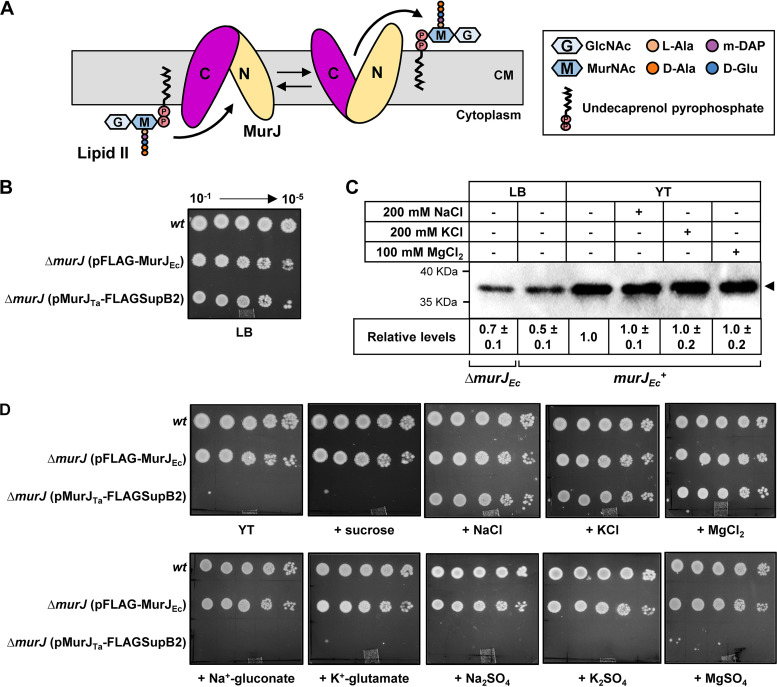
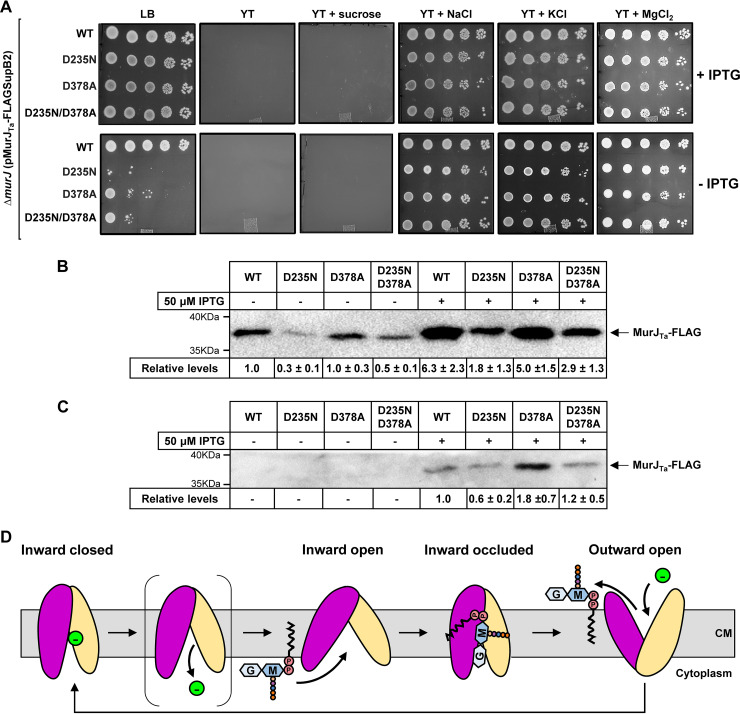
Most bacteria have a peptidoglycan cell wall that determines their cell shape and helps them resist osmotic lysis. Peptidoglycan synthesis depends on the translocation of the lipid-linked precursor lipid II across the cytoplasmic membrane by the MurJ flippase. Structure-function analyses of MurJ from Thermosipho africanus (MurJ) and Escherichia coli (MurJ) have revealed that MurJ adopts multiple conformations and utilizes an alternating-access mechanism to flip lipid II. MurJ activity relies on membrane potential, but the specific counterion has not been identified. Crystal structures of MurJ revealed a chloride ion bound to the N-lobe of the flippase and a sodium ion in its C-lobe, but the role of these ions in transport is unknown. Here, we investigated the effect of various ions on the function of MurJ and MurJ . We found that chloride, and not sodium, ions are necessary for MurJ function, but neither ion is required for MurJ function. We also showed that alleles encoding changes at the crystallographically identified sodium-binding site still complement the loss of native , although they decreased protein stability and/or function. Based on our data and previous work, we propose that chloride ions are necessary for the conformational change that resets MurJ after lipid II translocation and suggest that MurJ orthologs may function similarly but differ in their requirements for counterions. The biosynthetic pathway of the peptidoglycan cell wall is one of the most favorable targets for antibiotic development. Lipid II, the lipid-linked PG precursor, is made in the inner leaflet of the cytoplasmic membrane and then transported by the MurJ flippase so that it can be used to build the peptidoglycan cell wall. MurJ functions using an alternating-access mechanism thought to depend on a yet-to-be-identified counterion. This study fills a gap in our understanding of MurJ's energy-coupling mechanism by showing that chloride ions are required for MurJ in some, but not all, organisms. Based on our data and prior studies, we propose that, while the general transport mechanism of MurJ may be conserved, its specific mechanistic details may differ across bacteria, as is common in transporters. These findings are important to understand MurJ function and its development as an antibiotic target.
大多数细菌都有一层肽聚糖细胞壁,这层细胞壁决定了它们的细胞形状,并帮助它们抵抗渗透压裂解。肽聚糖的合成依赖于 MurJ 翻转酶将脂质连接前体脂质 II 穿过细胞质膜的易位。来自非洲嗜热泉古生菌(MurJ)和大肠杆菌(MurJ)的 MurJ 的结构-功能分析表明,MurJ 采用多种构象,并利用交替访问机制翻转脂质 II。MurJ 的活性依赖于膜电位,但具体的抗衡离子尚未确定。MurJ 的晶体结构显示,一个氯离子结合在翻转酶的 N 结构域,一个钠离子结合在 C 结构域,但这些离子在运输中的作用尚不清楚。在这里,我们研究了各种离子对 MurJ 和 MurJ 功能的影响。我们发现氯离子,而不是钠离子,是 MurJ 功能所必需的,但钠离子不是 MurJ 功能所必需的。我们还表明,晶体结构中鉴定的钠离子结合位点的 等位基因编码的变化仍然可以弥补天然 的缺失,尽管它们降低了蛋白质的稳定性和/或功能。基于我们的数据和以前的工作,我们提出氯离子是 MurJ 在后脂质 II 易位时重置构象变化所必需的,并且表明 MurJ 同源物可能具有类似的功能,但对抗衡离子的要求不同。 肽聚糖细胞壁的生物合成途径是抗生素开发最有希望的靶点之一。脂质 II,即肽聚糖前体的脂质连接物,在内质网膜的内层生成,然后由 MurJ 翻转酶转运,以便用于构建肽聚糖细胞壁。MurJ 利用一种交替访问机制发挥作用,这种机制被认为依赖于尚未确定的抗衡离子。这项研究通过表明氯离子是某些(但不是所有)生物体中 MurJ 的必需物质,填补了我们对 MurJ 能量偶联机制理解的空白。基于我们的数据和以前的研究,我们提出,虽然 MurJ 的一般转运机制可能是保守的,但它的具体机制细节可能因细菌而异,这在转运体中很常见。这些发现对于理解 MurJ 的功能及其作为抗生素靶标的开发非常重要。