Department of Computational Biology and Medical Sciences, Graduate School of Frontier Sciences, The University of Tokyo, Kashiwa, Chiba 277-8562, Japan.
Department of Biochemistry, Graduate School of Medicine, University of the Ryukyus, Nishihara-cho, Okinawa 903-0125, Japan.
Nucleic Acids Res. 2023 Mar 21;51(5):2434-2446. doi: 10.1093/nar/gkad081.
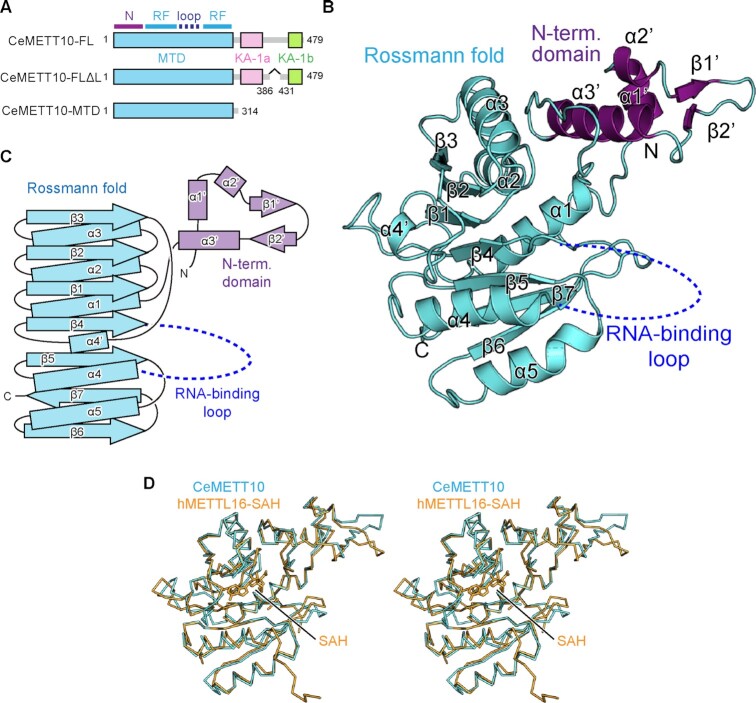
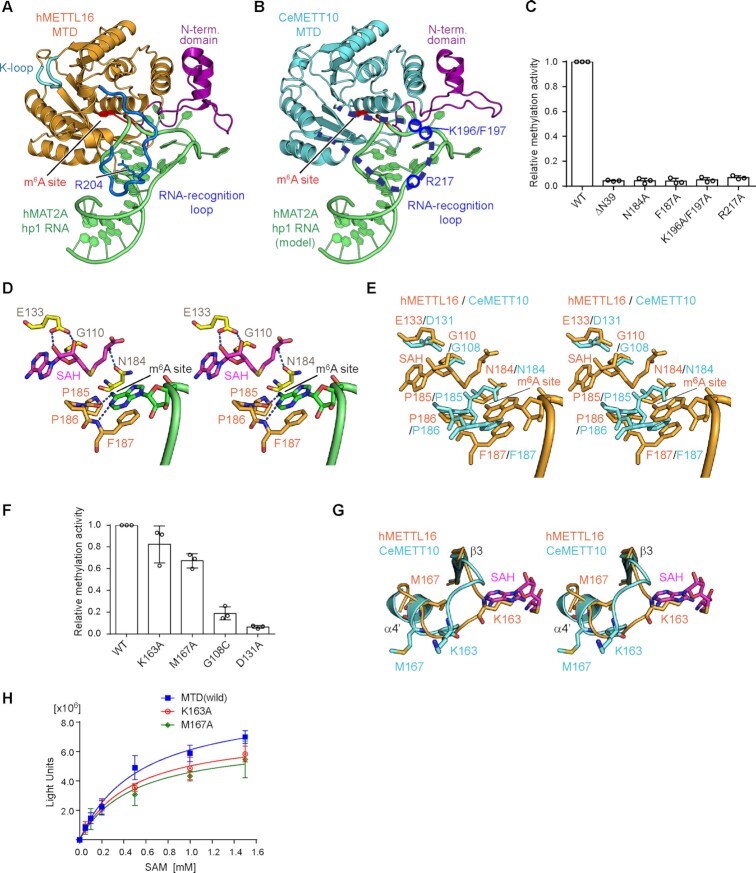
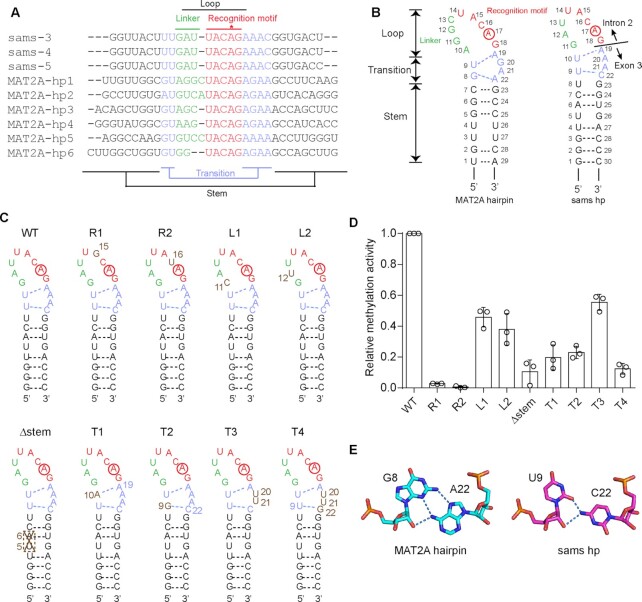
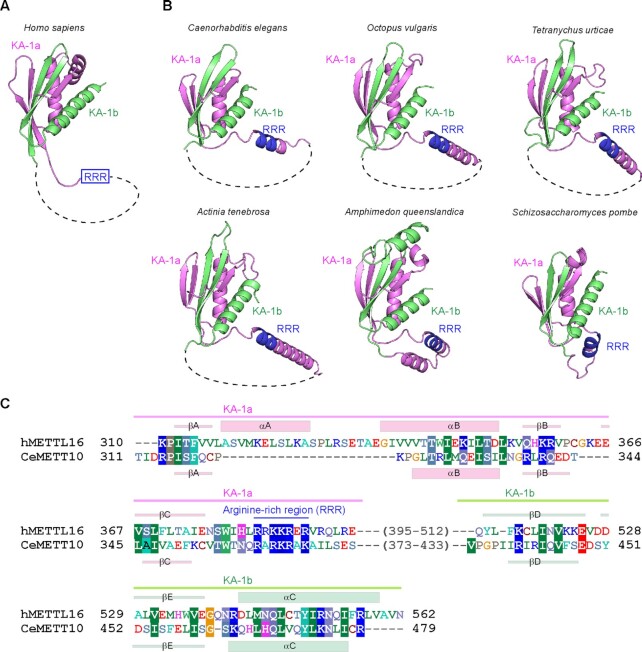
In Caenorhabditis elegans, the N6-methyladenosine (m6A) modification by METT10, at the 3'-splice sites in S-adenosyl-l-methionine (SAM) synthetase (sams) precursor mRNA (pre-mRNA), inhibits sams pre-mRNA splicing, promotes alternative splicing coupled with nonsense-mediated decay of the pre-mRNAs, and thereby maintains the cellular SAM level. Here, we present structural and functional analyses of C. elegans METT10. The structure of the N-terminal methyltransferase domain of METT10 is homologous to that of human METTL16, which installs the m6A modification in the 3'-UTR hairpins of methionine adenosyltransferase (MAT2A) pre-mRNA and regulates the MAT2A pre-mRNA splicing/stability and SAM homeostasis. Our biochemical analysis suggested that C. elegans METT10 recognizes the specific structural features of RNA surrounding the 3'-splice sites of sams pre-mRNAs, and shares a similar substrate RNA recognition mechanism with human METTL16. C. elegans METT10 also possesses a previously unrecognized functional C-terminal RNA-binding domain, kinase associated 1 (KA-1), which corresponds to the vertebrate-conserved region (VCR) of human METTL16. As in human METTL16, the KA-1 domain of C. elegans METT10 facilitates the m6A modification of the 3'-splice sites of sams pre-mRNAs. These results suggest the well-conserved mechanisms for the m6A modification of substrate RNAs between Homo sapiens and C. elegans, despite their different regulation mechanisms for SAM homeostasis.
在秀丽隐杆线虫中,METT10 在 S-腺苷甲硫氨酸(SAM)合成酶(sams)前体 mRNA(pre-mRNA)的 3'-剪接位点上将 N6-甲基腺苷(m6A)修饰,抑制 sams pre-mRNA 的剪接,促进与无意义介导的衰变偶联的选择性剪接前体 mRNA,并由此维持细胞 SAM 水平。在这里,我们对秀丽隐杆线虫 METT10 进行了结构和功能分析。METT10 的 N 端甲基转移酶结构域的结构与人类 METTL16 同源,后者在甲硫氨酸腺苷转移酶(MAT2A)pre-mRNA 的 3'-UTR 发夹中安装 m6A 修饰,并调节 MAT2A pre-mRNA 的剪接/稳定性和 SAM 稳态。我们的生化分析表明,秀丽隐杆线虫 METT10 识别 sams pre-mRNAs 3'-剪接位点周围 RNA 的特定结构特征,并与人类 METTL16 具有相似的底物 RNA 识别机制。秀丽隐杆线虫 METT10 还具有以前未被识别的功能 C 端 RNA 结合结构域激酶相关蛋白 1(KA-1),它对应于人类 METTL16 的脊椎动物保守区(VCR)。与人类 METTL16 一样,秀丽隐杆线虫 METT10 的 KA-1 结构域促进了 sams pre-mRNAs 3'-剪接位点的 m6A 修饰。这些结果表明,尽管人类和秀丽隐杆线虫的 SAM 稳态调节机制不同,但它们的底物 RNA m6A 修饰机制是高度保守的。