Cellular Biochemistry, Max-Planck-Institute for Multidisciplinary Sciences, Am Fassberg 11, Göttingen 37077, Germany.
Institut für Chemie und Biochemie, RNA Biochemie, Freie Universität Berlin, Takustr. 6, Berlin 14195, Germany.
Sci Adv. 2023 Mar 3;9(9):eadf1785. doi: 10.1126/sciadv.adf1785.
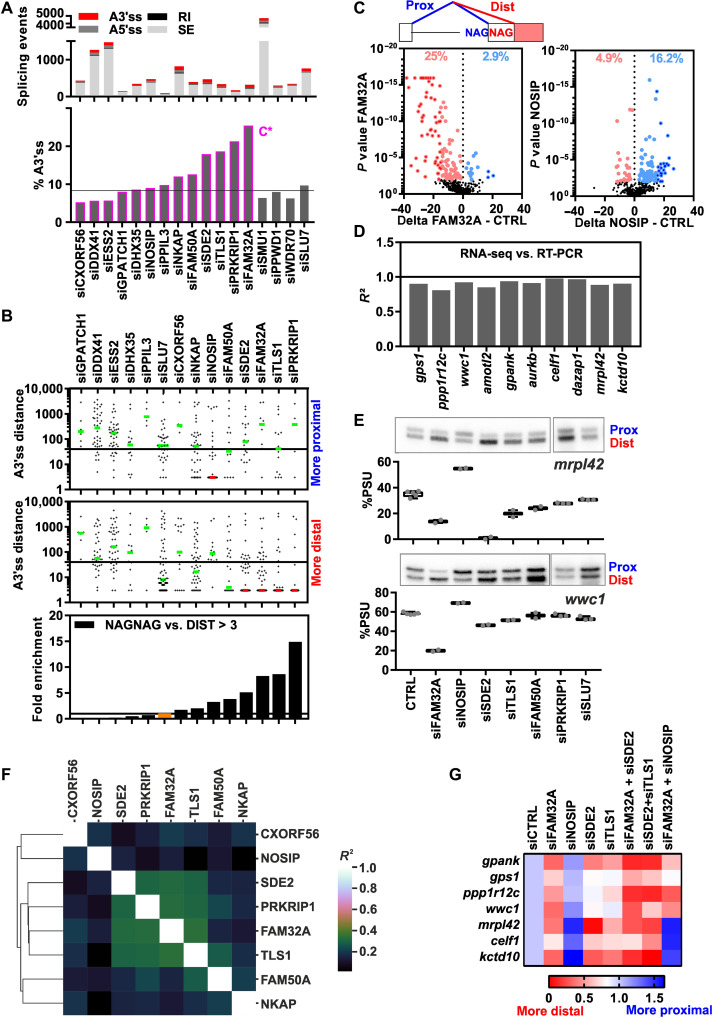
Alternative precursor messenger RNA splicing is instrumental in expanding the proteome of higher eukaryotes, and changes in 3' splice site (3'ss) usage contribute to human disease. We demonstrate by small interfering RNA-mediated knockdowns, followed by RNA sequencing, that many proteins first recruited to human C* spliceosomes, which catalyze step 2 of splicing, regulate alternative splicing, including the selection of alternatively spliced NAGNAG 3'ss. Cryo-electron microscopy and protein cross-linking reveal the molecular architecture of these proteins in C* spliceosomes, providing mechanistic and structural insights into how they influence 3'ss usage. They further elucidate the path of the 3' region of the intron, allowing a structure-based model for how the C* spliceosome potentially scans for the proximal 3'ss. By combining biochemical and structural approaches with genome-wide functional analyses, our studies reveal widespread regulation of alternative 3'ss usage after step 1 of splicing and the likely mechanisms whereby C* proteins influence NAGNAG 3'ss choices.
替代前体信使 RNA 剪接对于扩大高等真核生物的蛋白质组至关重要,而 3' 剪接位点 (3'ss) 使用的变化导致了人类疾病。我们通过小干扰 RNA 介导的敲低,随后进行 RNA 测序,证明许多最初被招募到人类 C* 剪接体的蛋白质,这些蛋白质可以催化剪接的第二步,调节选择性剪接,包括选择选择性剪接的 NAGNAG 3'ss。低温电子显微镜和蛋白质交联揭示了这些蛋白质在 C* 剪接体中的分子结构,为它们如何影响 3'ss 使用提供了机制和结构上的见解。它们进一步阐明了内含子 3' 区域的路径,为 C* 剪接体如何潜在地扫描近端 3'ss 提供了一个基于结构的模型。通过将生化和结构方法与全基因组功能分析相结合,我们的研究揭示了剪接第一步后广泛调节选择性 3'ss 使用的情况,以及 C* 蛋白影响 NAGNAG 3'ss 选择的可能机制。