Department of Pharmacology, Stony Brook University Renaissance School of Medicine, Stony Brook, NY 11794-8651, USA.
Host Pathogen Interaction Program, Southwest National Primate Research Center, Texas Biomedical Research Institute, San Antonio, TX 78227-5302, USA.
Viruses. 2023 Feb 24;15(3):623. doi: 10.3390/v15030623.
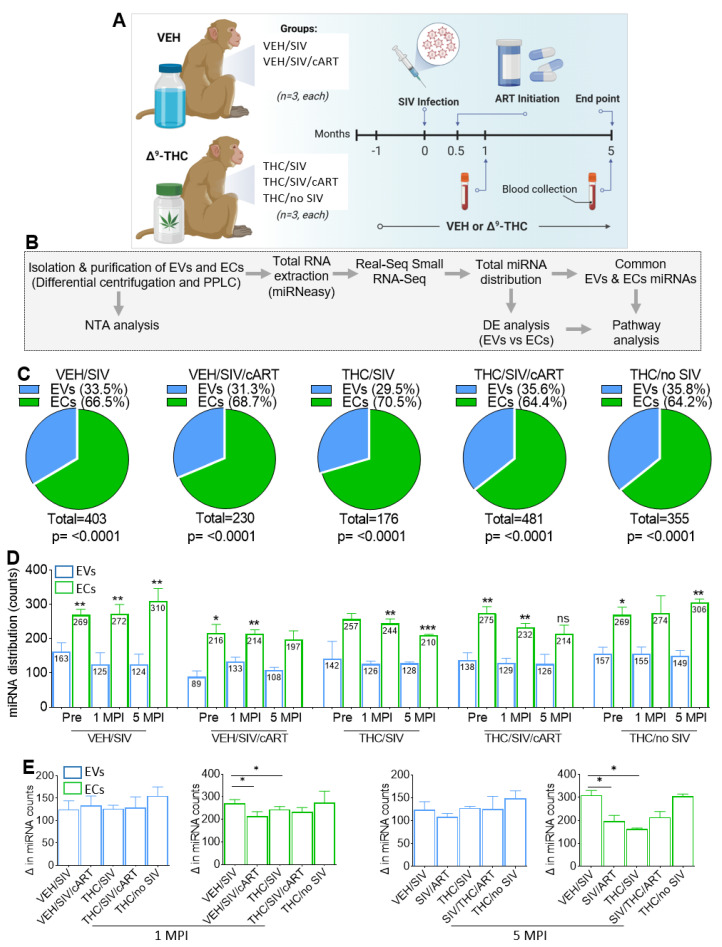
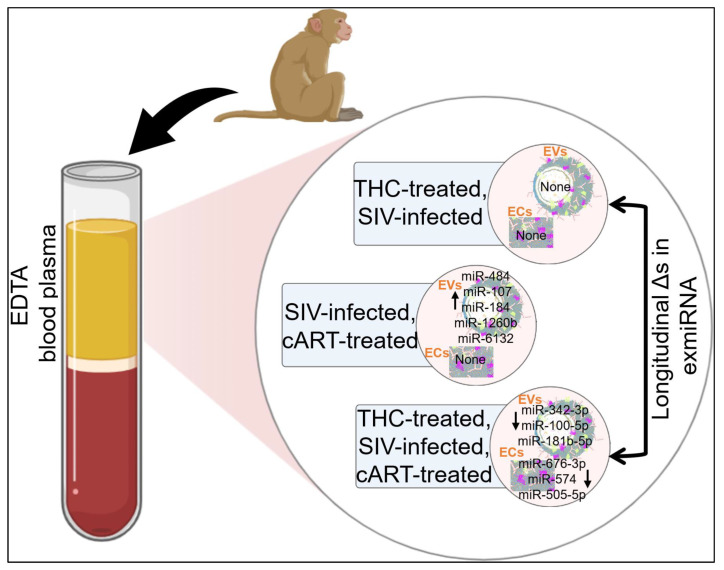
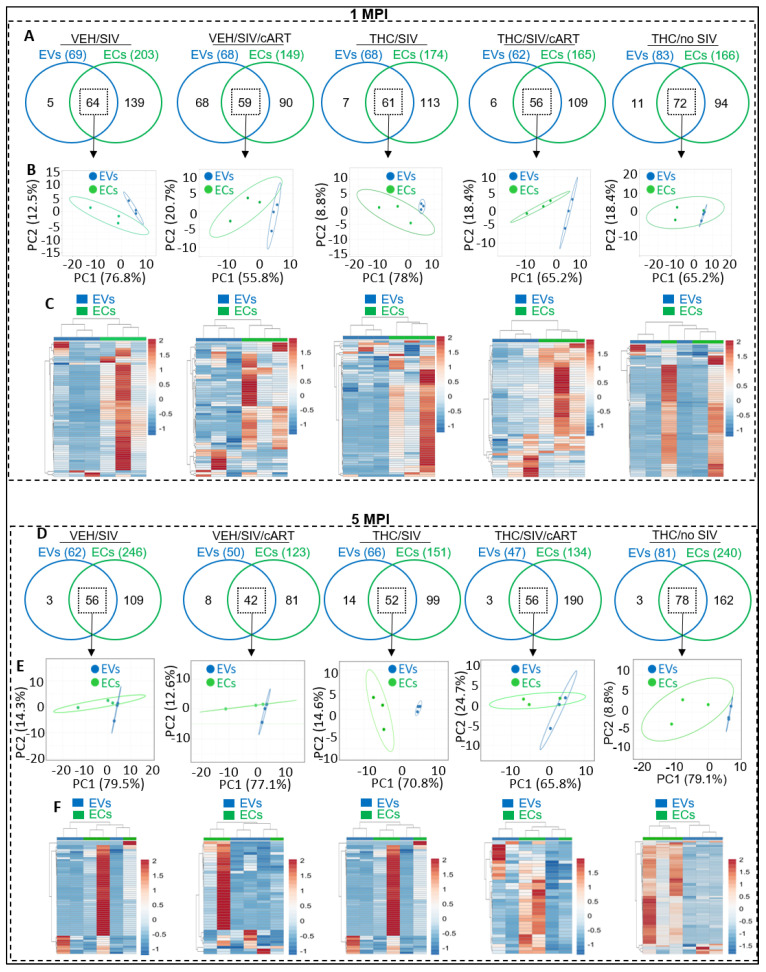
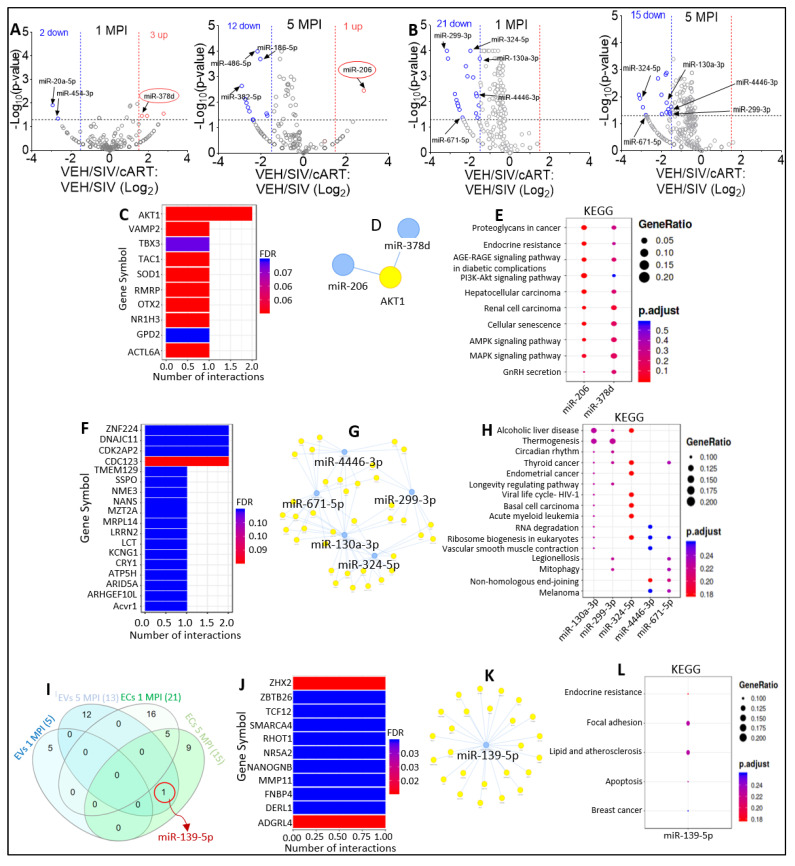
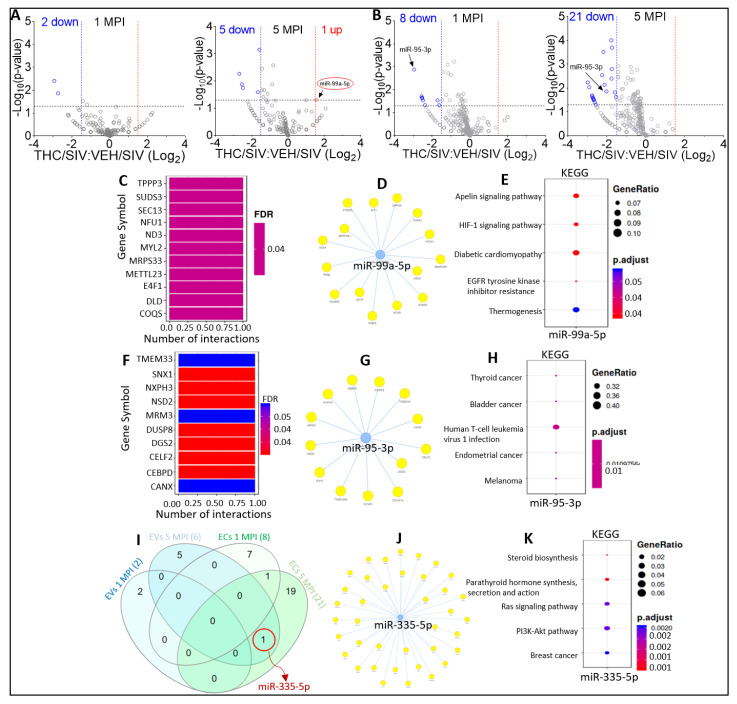
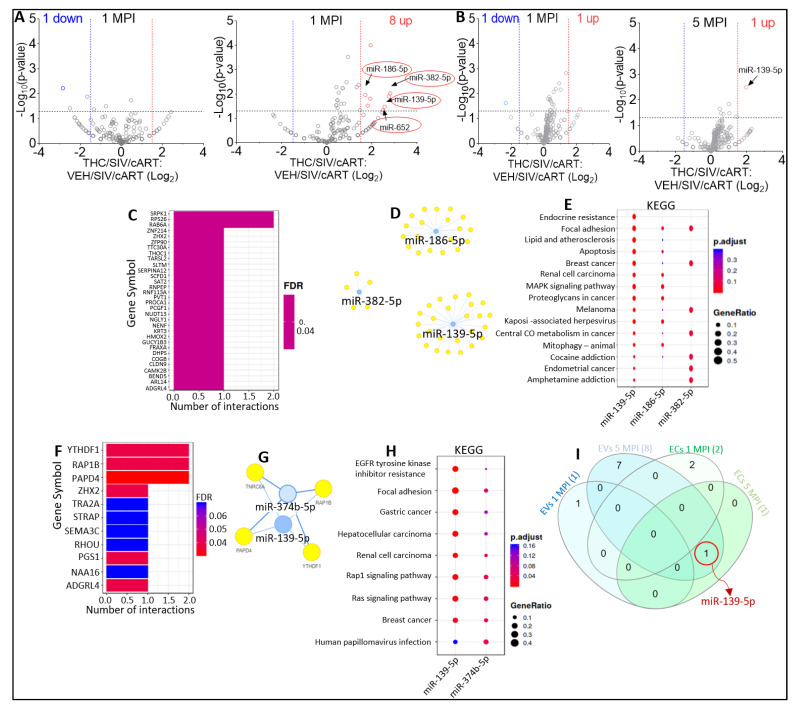
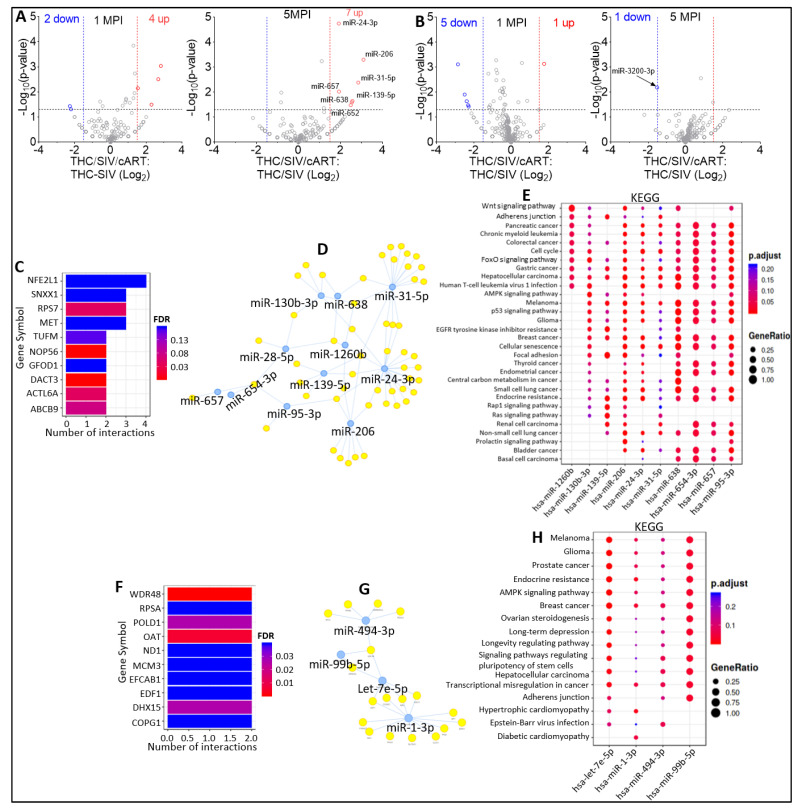
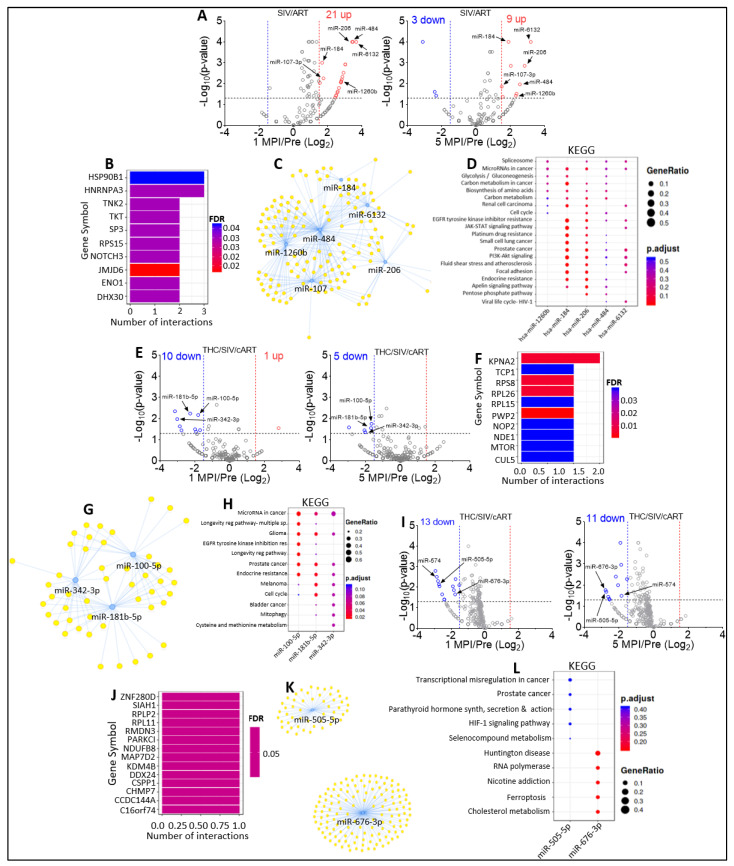
In this follow-up study, we investigated the abundance and compartmentalization of blood plasma extracellular miRNA (exmiRNA) into lipid-based carriers-blood plasma extracellular vesicles (EVs) and non-lipid-based carriers-extracellular condensates (ECs) during SIV infection. We also assessed how combination antiretroviral therapy (cART), administered in conjunction with phytocannabinoid delta-9-tetrahydrocannabinol (THC), altered the abundance and compartmentalization of exmiRNAs in the EVs and ECs of SIV-infected rhesus macaques (RMs). Unlike cellular miRNAs, exmiRNAs in blood plasma may serve as minimally invasive disease indicators because they are readily detected in stable forms. The stability of exmiRNAs in cell culture fluids and body fluids (urine, saliva, tears, cerebrospinal fluid (CSF), semen, blood) is based on their association with different carriers (lipoproteins, EVs, and ECs) that protect them from the activities of endogenous RNases. Here, we showed that in the blood plasma of uninfected control RMs, significantly less exmiRNAs were associated with EVs compared to the level (30% higher) associated with ECs, and that SIV infection altered the profile of EVs and ECs miRNAome (Manuscript 1). In people living with HIV (PLWH), host-encoded miRNAs regulate both host and viral gene expression, which may serve as indicators of disease or treatment biomarkers. The profile of miRNAs in blood plasma of PLWH (elite controllers versus viremic patients) are different, indicating that HIV may alter host miRNAome. However, there are no studies assessing the effect of cART or other substances used by PLWH, such as THC, on the abundance of exmiRNA and their association with EVs and ECs. Moreover, longitudinal exmiRNA profiles following SIV infection, treatment with THC, cART, or THC+cART remains unclear. Here, we serially analyzed miRNAs associated with blood plasma derived EVs and ECs. Paired EVs and ECs were separated from EDTA blood plasma of male Indian rhesus macaques (RMs) in five treatment groups, including VEH/SIV, VEH/SIV/cART, THC/SIV, THC/SIV/cART, or THC alone. Separation of EVs and ECs was achieved with the unparalleled nano-particle purification tool ─PPLC, a state-of-the-art, innovative technology equipped with gradient agarose bead sizes and a fast fraction collector that allows high resolution separation and retrieval of preparative quantities of sub-populations of extracellular structures. Global miRNA profiles of the paired EVs and ECs were determined with RealSeq Biosciences (Santa Cruz, CA) custom sequencing platform by conducting small RNA (sRNA)-seq. The sRNA-seq data were analyzed using various bioinformatic tools. Validation of key exmiRNA was performed using specific TaqMan microRNA stem-loop RT-qPCR assays. We investigated the effect of cART, THC, or both cART and THC together on the abundance and compartmentalization of blood plasma exmiRNA in EVs and ECs in SIV-infected RMs. As shown in Manuscript 1 of this series, were in uninfected RMs, ~30% of exmiRNAs were associated with ECs, we confirmed in this follow up manuscript that exmiRNAs were present in both lipid-based carriers-EVs and non-lipid-based carriers-ECs, with 29.5 to 35.6% and 64.2 to 70.5 % being associated with EVs and ECs, respectively. Remarkably, the different treatments (cART, THC) have distinct effects on the enrichment and compartmentalization pattern of exmiRNAs. In the VEH/SIV/cART group, 12 EV-associated and 15 EC-associated miRNAs were significantly downregulated. EV-associated miR-206, a muscle-specific miRNA that is present in blood, was higher in the VEH/SIV/ART compared to the VEH/SIV group. ExmiR-139-5p that was implicated in endocrine resistance, focal adhesion, lipid and atherosclerosis, apoptosis, and breast cancer by miRNA-target enrichment analysis was significantly lower in VEH/SIV/cART compared to VEH/SIV, irrespective of the compartment. With respect to THC treatment, 5 EV-associated and 21 EC-associated miRNAs were significantly lower in the VEH/THC/SIV. EV-associated miR-99a-5p was higher in VEH/THC/SIV compared to VEH/SIV, while miR-335-5p counts were significantly lower in both EVs and ECs of THC/SIV compared to VEH/SIV. EVs from SIV/cART/THC combined treatment group have significant increases in the count of eight (miR-186-5p, miR-382-5p, miR-139-5p and miR-652, miR-10a-5p, miR-657, miR-140-5p, miR-29c-3p) miRNAs, all of which were lower in VEH/SIV/cART group. Analysis of miRNA-target enrichment showed that this set of eight miRNAs were implicated in endocrine resistance, focal adhesions, lipid and atherosclerosis, apoptosis, and breast cancer as well as cocaine and amphetamine addiction. In ECs and EVs, combined THC and cART treatment significantly increased miR-139-5p counts compared to VEH/SIV group. Significant alterations in these host miRNAs in both EVs and ECs in the untreated and treated (cART, THC, or both) RMs indicate the persistence of host responses to infection or treatments, and this is despite cART suppression of viral load and THC suppression of inflammation. To gain further insight into the pattern of miRNA alterations in EVs and ECs and to assess potential cause-and-effect relationships, we performed longitudinal miRNA profile analysis, measured in terms of months (1 and 5) post-infection (MPI). We uncovered miRNA signatures associated with THC or cART treatment of SIV-infected macaques in both EVs and ECs. While the number of miRNAs was significantly higher in ECs relative to EVs for all groups (VEH/SIV, SIV/cART, THC/SIV, THC/SIV/cART, and THC) longitudinally from 1 MPI to 5 MPI, treatment with cART and THC have longitudinal effects on the abundance and compartmentalization pattern of exmiRNAs in the two carriers. As shown in Manuscript 1 where SIV infection led to longitudinal suppression of EV-associated miRNA-128-3p, administration of cART to SIV-infected RMs did not increase miR-128-3p but resulted in longitudinal increases in six EV-associated miRNAs (miR-484, miR-107, miR-206, miR-184, miR-1260b, miR-6132). Furthermore, administration of cART to THC treated SIV-infected RMs resulted in a longitudinal decrease in three EV-associated miRNAs (miR-342-3p, miR-100-5p, miR181b-5p) and a longitudinal increase in three EC-associated miRNAs (miR-676-3p, miR-574-3p, miR-505-5p). The longitudinally altered miRNAs in SIV-infected RMs may indicate disease progression, while in the cART Group and the THC Group, the longitudinally altered miRNAs may serve as biomarkers of response to treatment. This paired EVs and ECs miRNAome analyses provided a comprehensive cross-sectional and longitudinal summary of the host exmiRNA responses to SIV infection and the impact of THC, cART, or THC and cART together on the miRNAome during SIV infection. Overall, our data point to previously unrecognized alterations in the exmiRNA profile in blood plasma following SIV infection. Our data also indicate that cART and THC treatment independently and in combination may alter both the abundance and the compartmentalization of several exmiRNA related to various disease and biological processes.
在这项后续研究中,我们调查了在 SIV 感染期间,血浆细胞外 miRNA(exmiRNA)进入脂质载体-血浆细胞外囊泡(EVs)和非脂质载体-细胞外凝聚物(ECs)的丰度和区室化。我们还评估了联合抗逆转录病毒疗法(cART)与植物大麻素 delta-9-四氢大麻酚(THC)一起给药如何改变 SIV 感染的食蟹猴(RMs)血浆 EVs 和 ECs 中外泌 miRNA 的丰度和区室化。与细胞内 miRNA 不同,血浆中外泌 miRNA 可能作为微创疾病指标,因为它们以稳定的形式容易被检测到。exmiRNA 在细胞培养液和体液(尿液、唾液、眼泪、脑脊液(CSF)、精液、血液)中的稳定性基于它们与不同载体(脂蛋白、EVs 和 ECs)的关联,这些载体保护它们免受内源性 RNase 的影响。在这里,我们表明,在未感染对照 RM 的血浆中,与 EVs 相关的 exmiRNA 水平(比与 ECs 相关的水平高 30%)明显低于与 ECs 相关的水平,并且 SIV 感染改变了 EVs 和 ECs miRNAome 的分布(文献 1)。在 HIV 感染者(PLWH)中,宿主编码的 miRNA 调节宿主和病毒基因的表达,这可能作为疾病或治疗生物标志物的指标。PLWH 血浆中 miRNA 的特征(精英控制器与病毒载量患者)不同,表明 HIV 可能改变宿主 miRNAome。然而,目前尚无研究评估 cART 或 PLWH 等其他物质(如 THC)对 exmiRNA 的丰度及其与 EVs 和 ECs 的关联的影响。此外,SIV 感染、THC、cART 或 THC+cART 治疗后的血浆 exmiRNA 纵向谱尚不清楚。在这里,我们对从雄性印度恒河猴(RMs)的 EDTA 血浆中分离的 EVs 和 ECs 相关的 miRNA 进行了连续分析。在五个治疗组中,包括 VEH/SIV、VEH/SIV/cART、THC/SIV、THC/SIV/cART 和 THC 单独治疗组,分离 EVs 和 ECs。使用无与伦比的纳米粒子净化工具-PPLC 实现了 EVs 和 ECs 的分离,PPLC 是一种最先进的创新技术,配备了梯度琼脂糖珠大小和快速的馏分收集器,可实现亚群细胞外结构的高分辨率分离和回收。使用 RealSeq Biosciences(圣克鲁斯,CA)定制测序平台通过进行小 RNA(sRNA)-seq 确定了配对 EVs 和 ECs 的全局 miRNA 谱。使用各种生物信息学工具分析 sRNA-seq 数据。使用特定的 TaqMan 微 RNA 茎环 RT-qPCR 测定法对关键 exmiRNA 进行验证。我们研究了 cART、THC 或两者联合治疗对 SIV 感染 RMs 血浆 EVs 和 ECs 中外泌 miRNA 丰度和区室化的影响。如本系列中的文献 1 所示,在未感染的 RM 中,~30%的 exmiRNA 与 ECs 相关,我们在本后续手稿中证实,exmiRNA 存在于脂质载体-EVs 和非脂质载体-ECs 中,分别有 29.5%到 35.6%和 64.2%到 70.5%与 EVs 和 ECs 相关。值得注意的是,不同的治疗方法(cART、THC)对 exmiRNA 的富集和区室化模式有明显的影响。在 VEH/SIV/cART 组中,12 个 EV 相关和 15 个 EC 相关 miRNA 显著下调。EV 相关的 miR-206 是一种存在于血液中的肌肉特异性 miRNA,在 VEH/SIV/ART 组中高于 VEH/SIV 组。miR-139-5p 通过 miRNA 靶标富集分析被牵连到内分泌抵抗、焦点粘连、脂质和动脉粥样硬化、凋亡和乳腺癌,在 VEH/SIV/cART 组中与 VEH/SIV 相比显著降低,而与区室无关。至于 THC 治疗,VEH/THC/SIV 中 5 个 EV 相关和 21 个 EC 相关 miRNA 显著下调。VEH/THC/SIV 中 miR-99a-5p 高于 VEH/SIV,而 miR-335-5p 在 EVs 和 ECs 中的计数均显著低于 VEH/SIV。与 SIV/cART/THC 联合治疗组相比,EVs 的计数显著增加了八种(miR-186-5p、miR-382-5p、miR-139-5p 和 miR-652、miR-10a-5p、miR-657、miR-140-5p、miR-29c-3p)miRNA,所有这些在 VEH/SIV/cART 组中均较低。miRNA 靶标富集分析表明,这组八个 miRNA 与内分泌抵抗、焦点粘连、脂质和动脉粥样硬化、凋亡以及可卡因和安非他命成瘾有关。在 ECs 和 EVs 中,联合 THC 和 cART 治疗与 VEH/SIV 组相比,显著增加了 miR-139-5p 的计数。在未治疗和治疗(cART、THC 或两者)RM 中,EVs 和 ECs 中外泌 miRNA 的这些显著改变表明宿主对感染或治疗的反应持续存在,尽管 cART 抑制了病毒载量,而 THC 抑制了炎症。为了更深入地了解 EVs 和 ECs 中外泌 miRNA 改变的模式,并评估潜在的因果关系,我们进行了纵向 miRNA 谱分析,以月(1 个月和 5 个月)为单位进行测量(MPI)。我们发现了与 SIV 感染的猕猴在 EVs 和 ECs 中用 THC 或 cART 治疗相关的 miRNA 特征。虽然在所有组(VEH/SIV、SIV/cART、THC/SIV、THC/SIV/cART 和 THC)中,从 1 MPI 到 5 MPI 时,ECs 中外泌 miRNA 的数量相对于 EVs 显著更高,但 cART 和 THC 的治疗对 EVs 和 ECs 中两种载体中外泌 miRNA 的丰度和区室化模式具有纵向作用。如文献 1 所示,SIV 感染导致 EV 相关的 miR-128-3p 纵向抑制,cART 对 SIV 感染的 RM 的给药并没有增加 miR-128-3p,但导致六个 EV 相关 miRNA(miR-484、miR-107、miR-206、miR-184、miR-1260b 和 miR-6132)的纵向增加。此外,cART 给药到用 THC 治疗的 SIV 感染的 RM 中,导致三个 EV 相关 miRNA(miR-342-3p、miR-100-5p、miR181b-5p)的纵向减少和三个 EC 相关 miRNA(miR-676-3p、miR-574-3p、miR-505-5p)的纵向增加。SIV 感染的 RM 中纵向改变的 miRNA 可能表明疾病进展,而在 cART 组和 THC 组中,纵向改变的 miRNA 可能作为治疗反应的生物标志物。这项配对 EVs 和 ECs miRNAome 分析提供了对 SIV 感染和 THC、cART 或 THC 和 cART 联合治疗期间宿主 exmiRNA 反应的全面交叉和纵向总结,用于 SIV 感染。总的来说,我们的数据指出了之前未被识别的 SIV 感染后血浆中外泌 miRNA 谱的改变。我们的数据还表明,cART 和 THC 治疗独立和联合治疗可能改变与各种疾病和生物过程相关的多种 exmiRNA 的丰度和区室化。