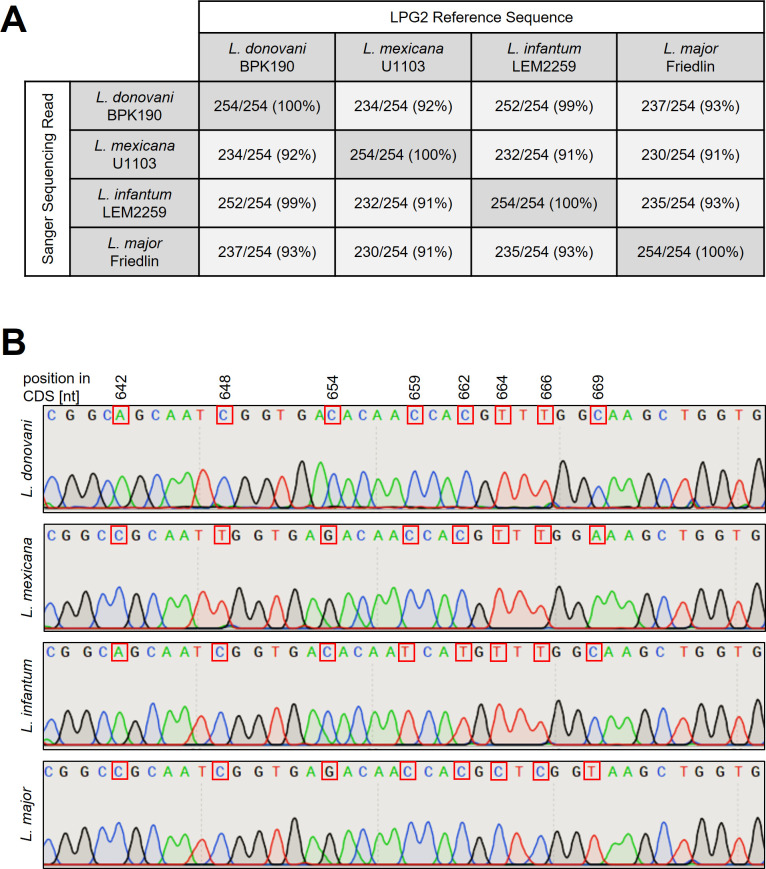
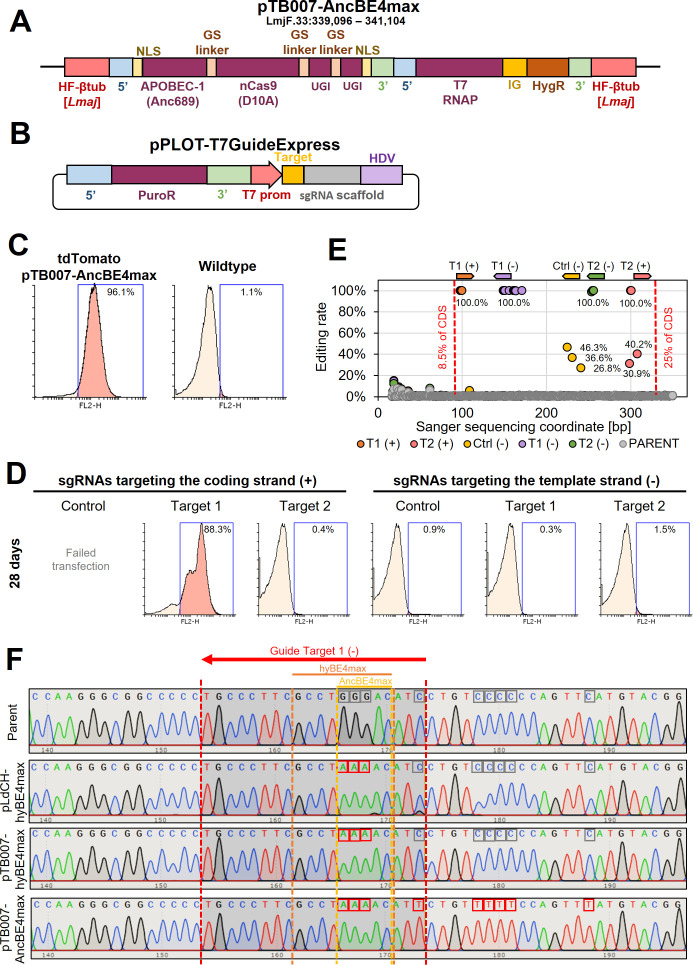
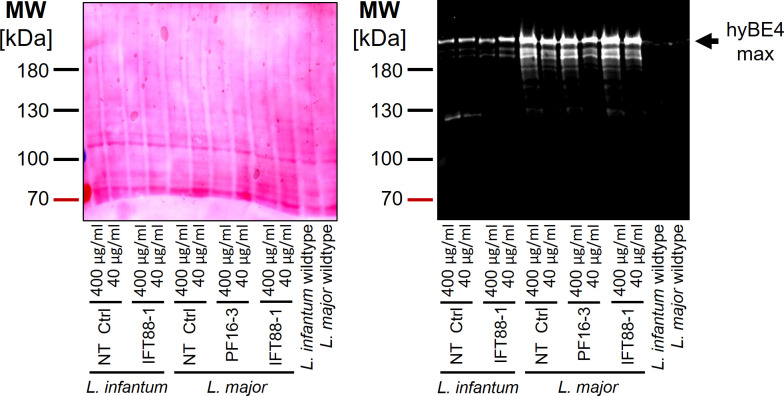
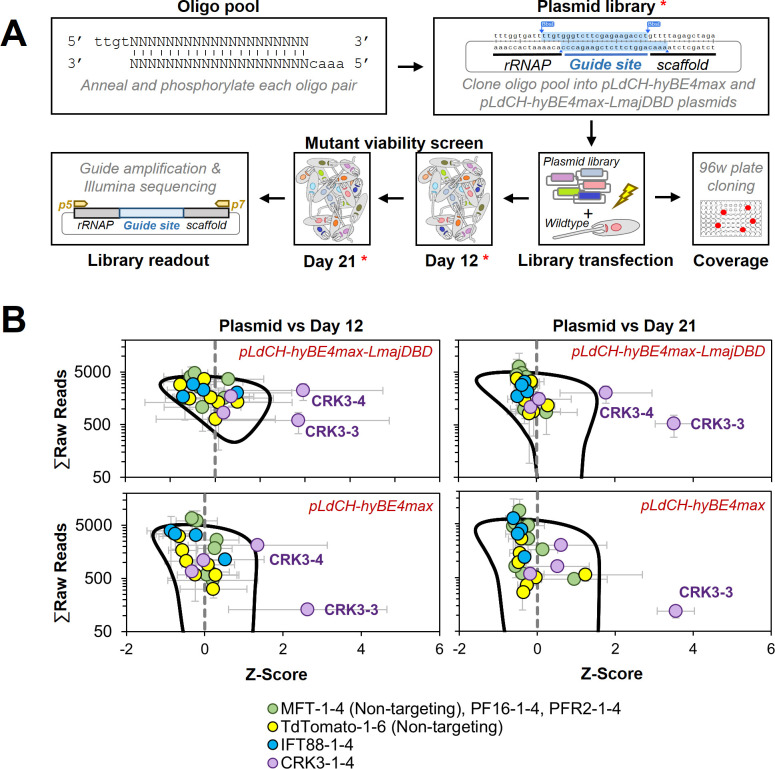
Department of Cell and Developmental Biology, Biocentre, University of Würzburg, Wuerzburg, Germany.
Elife. 2023 May 24;12:e85605. doi: 10.7554/eLife.85605.
CRISPR/Cas9 gene editing has revolutionised loss-of-function experiments in , the causative agent of leishmaniasis. As lack a functional non-homologous DNA end joining pathway however, obtaining null mutants typically requires additional donor DNA, selection of drug resistance-associated edits or time-consuming isolation of clones. Genome-wide loss-of-function screens across different conditions and across multiple species are therefore unfeasible at present. Here, we report a CRISPR/Cas9 cytosine base editor (CBE) toolbox that overcomes these limitations. We employed CBEs in to introduce STOP codons by converting cytosine into thymine and created http://www.leishbaseedit.net/ for CBE primer design in kinetoplastids. Through reporter assays and by targeting single- and multi-copy genes in , , , and , we demonstrate how this tool can efficiently generate functional null mutants by expressing just one single-guide RNA, reaching up to 100% editing rate in non-clonal populations. We then generated a -optimised CBE and successfully targeted an essential gene in a plasmid library delivered loss-of-function screen in . Since our method does not require DNA double-strand breaks, homologous recombination, donor DNA, or isolation of clones, we believe that this enables for the first time functional genetic screens in via delivery of plasmid libraries.
CRISPR/Cas9 基因编辑技术革新了 (引起利什曼病的病原体)的功能丧失实验。然而,由于 缺乏功能性的非同源 DNA 末端连接途径,因此通常需要额外的供体 DNA、选择与药物抗性相关的编辑或耗时的克隆分离,才能获得缺失突变体。目前,在不同条件下和多个 物种中进行全基因组功能丧失筛选是不可行的。在这里,我们报告了一个 CRISPR/Cas9 胞嘧啶碱基编辑器(CBE)工具包,该工具包克服了这些限制。我们在 中使用 CBE 将胞嘧啶转化为胸腺嘧啶,从而引入终止密码子,并创建了 http://www.leishbaseedit.net/ ,用于锥虫中 CBE 引物的设计。通过报告基因检测,并针对 、 、 和 中的单拷贝和多拷贝基因,我们展示了如何仅通过表达一个单向导 RNA 就可以有效地生成功能缺失突变体,在非克隆群体中达到高达 100%的编辑率。然后,我们生成了一个 - 优化的 CBE,并在 中成功靶向了质粒文库传递的功能丧失筛选中的一个必需基因。由于我们的方法不需要双链 DNA 断裂、同源重组、供体 DNA 或克隆分离,我们相信这首次使得通过质粒文库传递能够在 中进行功能遗传筛选。