Developmental Biology Program, Sloan-Kettering Institute, New York, New York, United States of America.
Laboratoire de Biologie et Modélisation de la Cellule, École Normale Supérieure de Lyon CNRS UMR5239, Université Claude Bernard Lyon 1, Lyon, France.
PLoS Biol. 2023 Jun 8;21(6):e3002136. doi: 10.1371/journal.pbio.3002136. eCollection 2023 Jun.
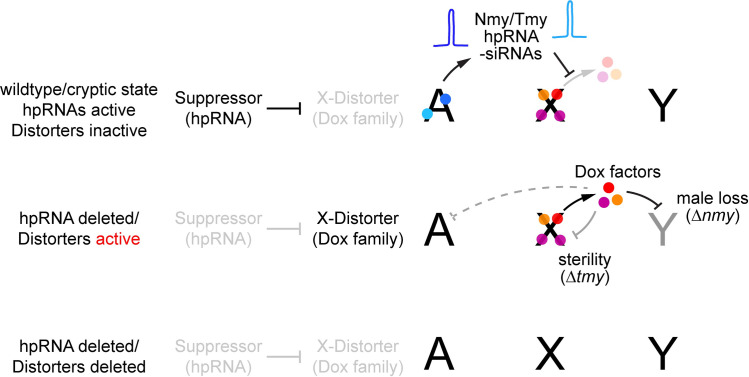
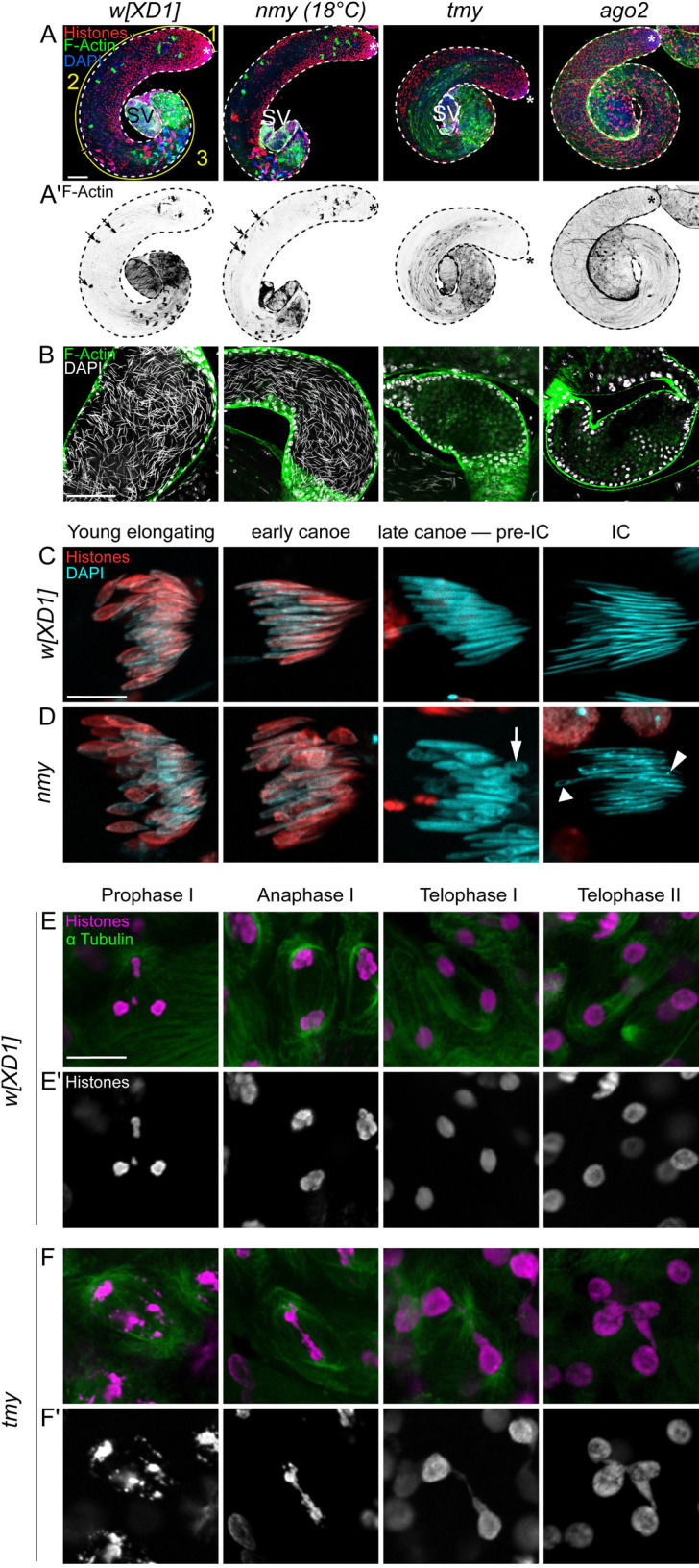
Meiotic drive loci distort the normally equal segregation of alleles, which benefits their own transmission even in the face of severe fitness costs to their host organism. However, relatively little is known about the molecular identity of meiotic drivers, their strategies of action, and mechanisms that can suppress their activity. Here, we present data from the fruitfly Drosophila simulans that address these questions. We show that a family of de novo, protamine-derived X-linked selfish genes (the Dox gene family) is silenced by a pair of newly emerged hairpin RNA (hpRNA) small interfering RNA (siRNA)-class loci, Nmy and Tmy. In the w[XD1] genetic background, knockout of nmy derepresses Dox and MDox in testes and depletes male progeny, whereas knockout of tmy causes misexpression of PDox genes and renders males sterile. Importantly, genetic interactions between nmy and tmy mutant alleles reveal that Tmy also specifically maintains male progeny for normal sex ratio. We show the Dox loci are functionally polymorphic within D. simulans, such that both nmy-associated sex ratio bias and tmy-associated sterility can be rescued by wild-type X chromosomes bearing natural deletions in different Dox family genes. Finally, using tagged transgenes of Dox and PDox2, we provide the first experimental evidence Dox family genes encode proteins that are strongly derepressed in cognate hpRNA mutants. Altogether, these studies support a model in which protamine-derived drivers and hpRNA suppressors drive repeated cycles of sex chromosome conflict and resolution that shape genome evolution and the genetic control of male gametogenesis.
减数分裂驱动基因座会扭曲等位基因的正常均等分离,即使对宿主生物有严重的适应代价,也有利于它们自身的传递。然而,关于减数分裂驱动基因的分子身份、它们的作用策略以及可以抑制其活性的机制,我们知之甚少。在这里,我们提供了来自果蝇 simulans 的数据,这些数据解决了这些问题。我们表明,一组新出现的发夹 RNA(hpRNA)小干扰 RNA(siRNA)类基因座 Nmy 和 Tmy 沉默了一组从头出现的、鱼精蛋白衍生的 X 连锁自私基因(Dox 基因家族)。在 w[XD1]遗传背景下,nmy 的敲除会在睾丸中解除 Dox 和 MDox 的抑制,导致雄性后代减少,而 tmy 的敲除会导致 PDox 基因的错误表达,并使雄性不育。重要的是,nmy 和 tmy 突变等位基因的遗传相互作用表明,Tmy 还专门为维持正常性别比例而维持雄性后代。我们表明,Dox 基因座在 D. simulans 内具有功能多态性,因此,nmy 相关的性别比例偏倚和 tmy 相关的不育都可以通过携带不同 Dox 家族基因天然缺失的野生型 X 染色体来挽救。最后,我们使用 Dox 和 PDox2 的标记转基因,提供了第一个实验证据,表明 Dox 家族基因编码的蛋白质在同源 hpRNA 突变体中强烈被解除抑制。总之,这些研究支持了一个模型,即鱼精蛋白衍生的驱动基因和 hpRNA 抑制剂驱动了性染色体冲突和解决的反复循环,从而塑造了基因组进化和雄性配子发生的遗传控制。