Institute of Neuroscience and Medicine INM-1, Research Centre Jülich, Jülich, Germany.
Center for Neural Science, New York University, New York, United States.
Elife. 2023 Aug 14;12:e82850. doi: 10.7554/eLife.82850.
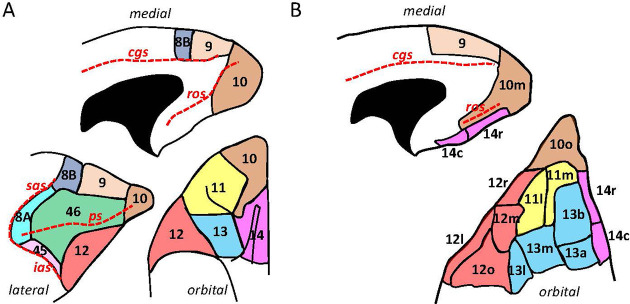
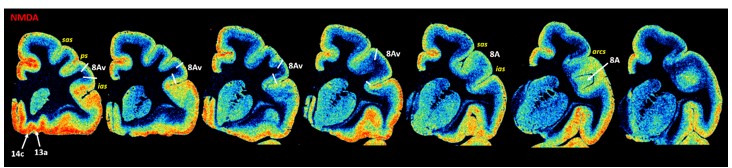
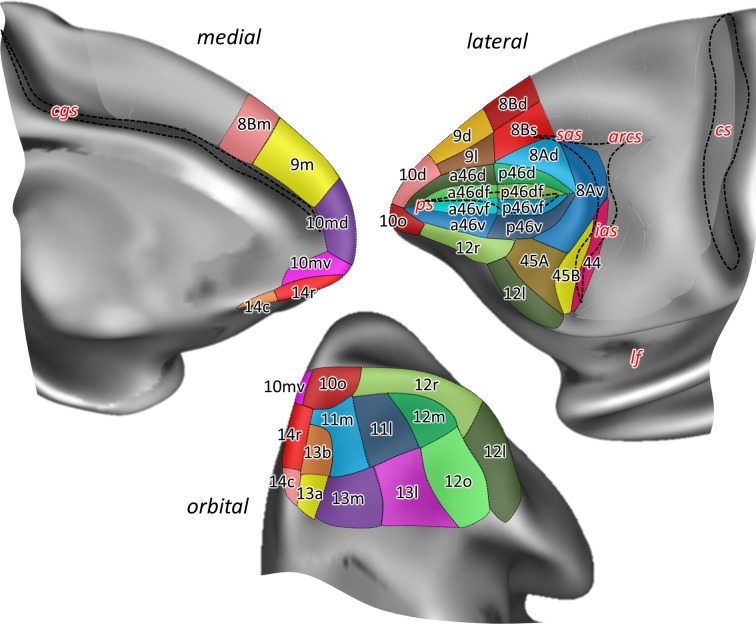
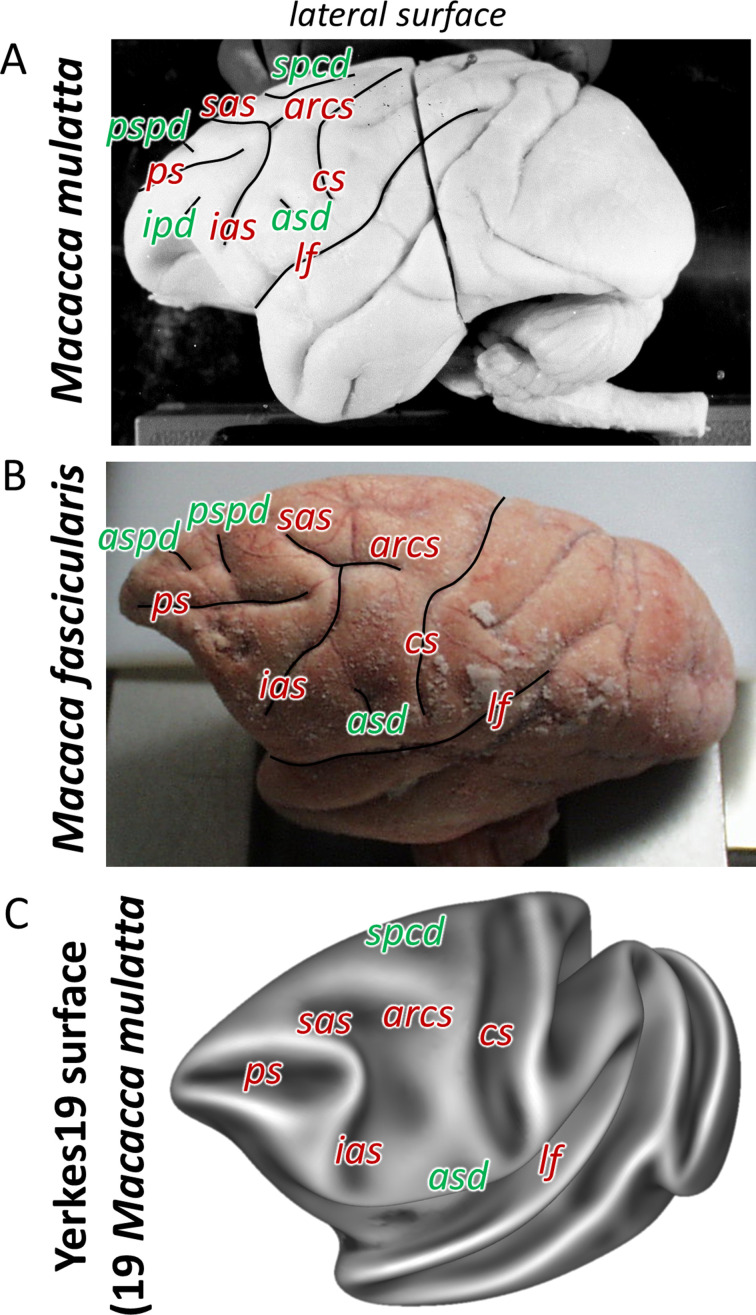
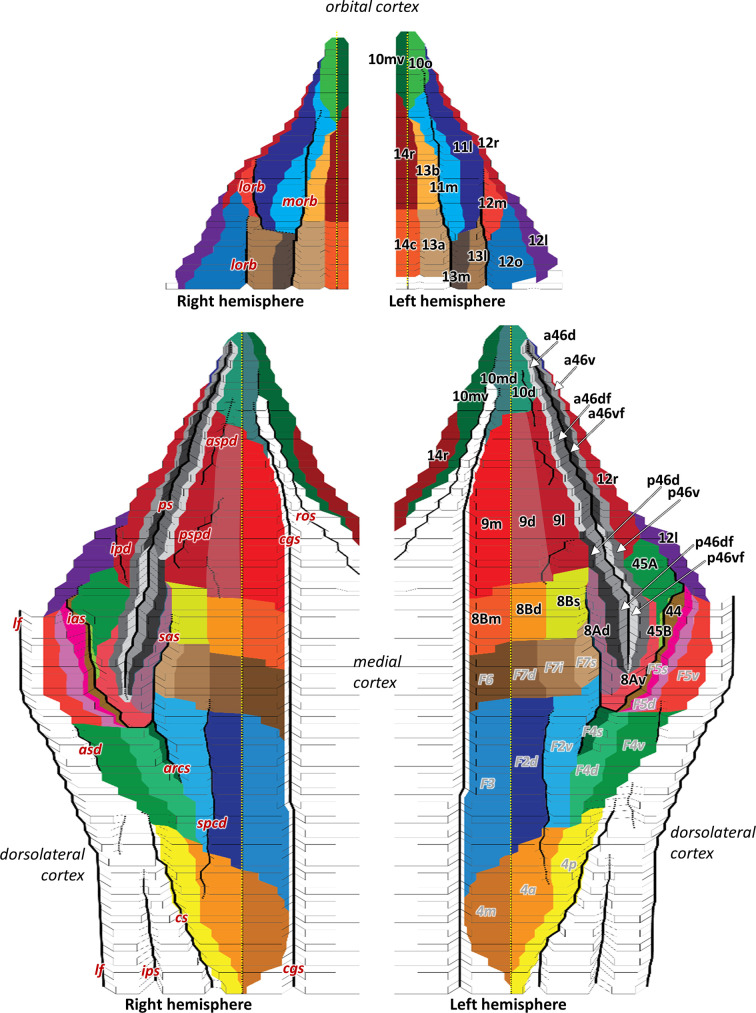
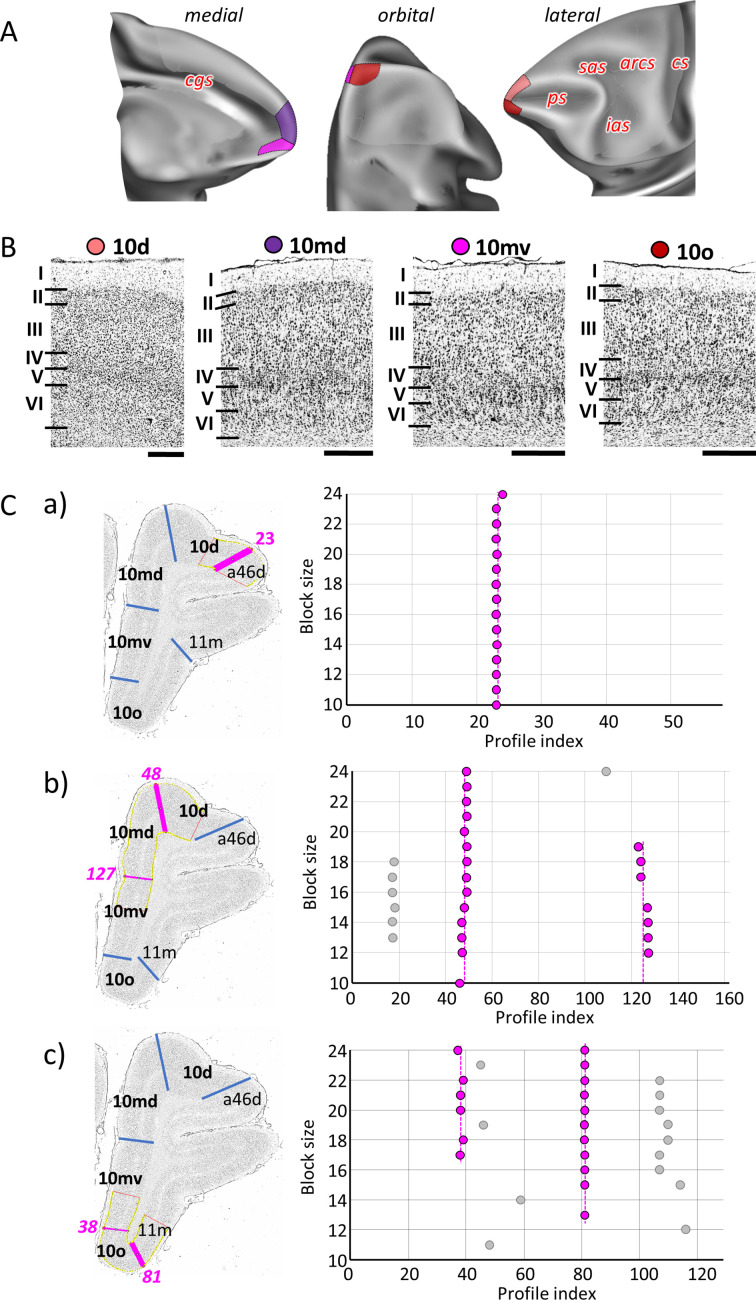
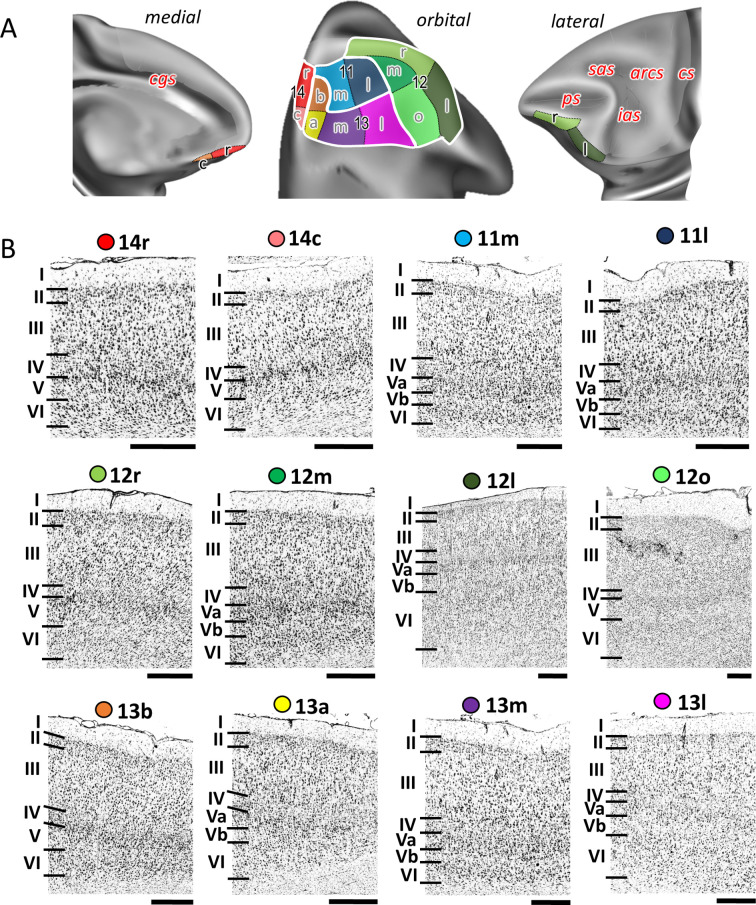
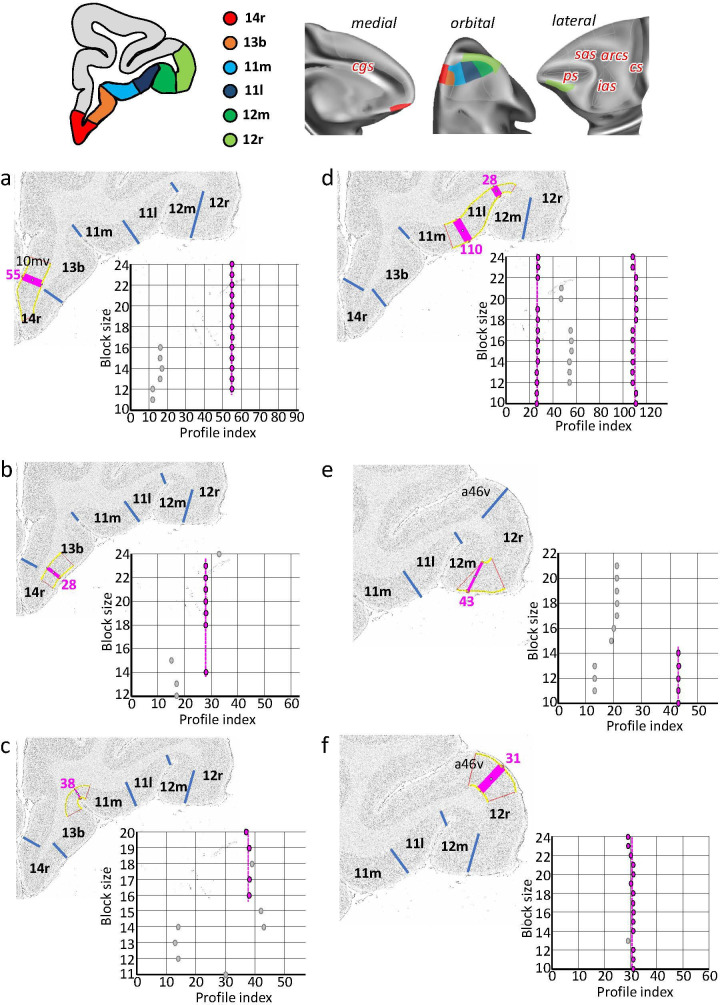
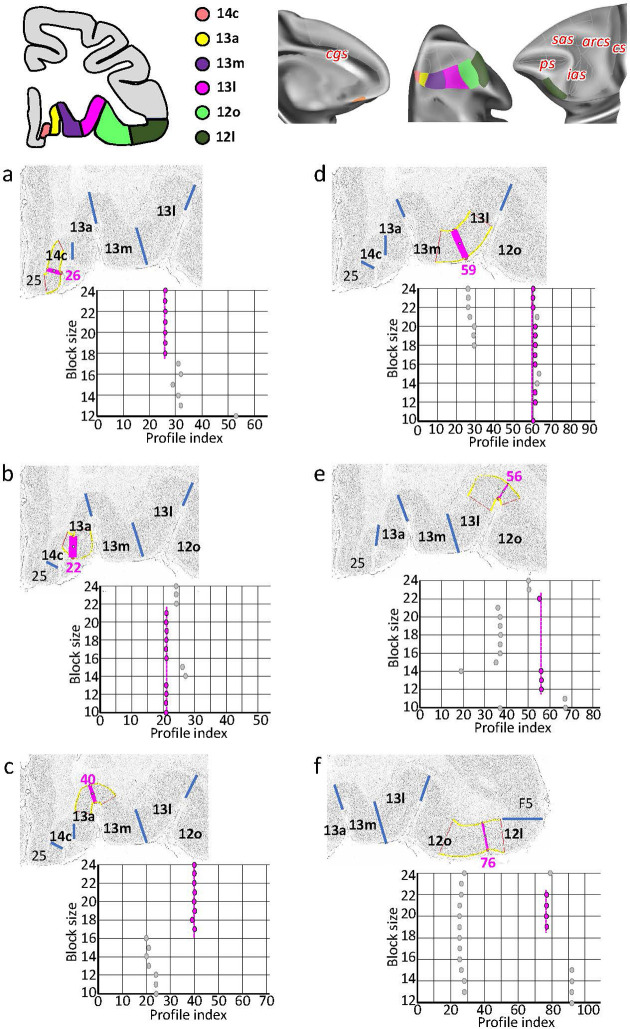
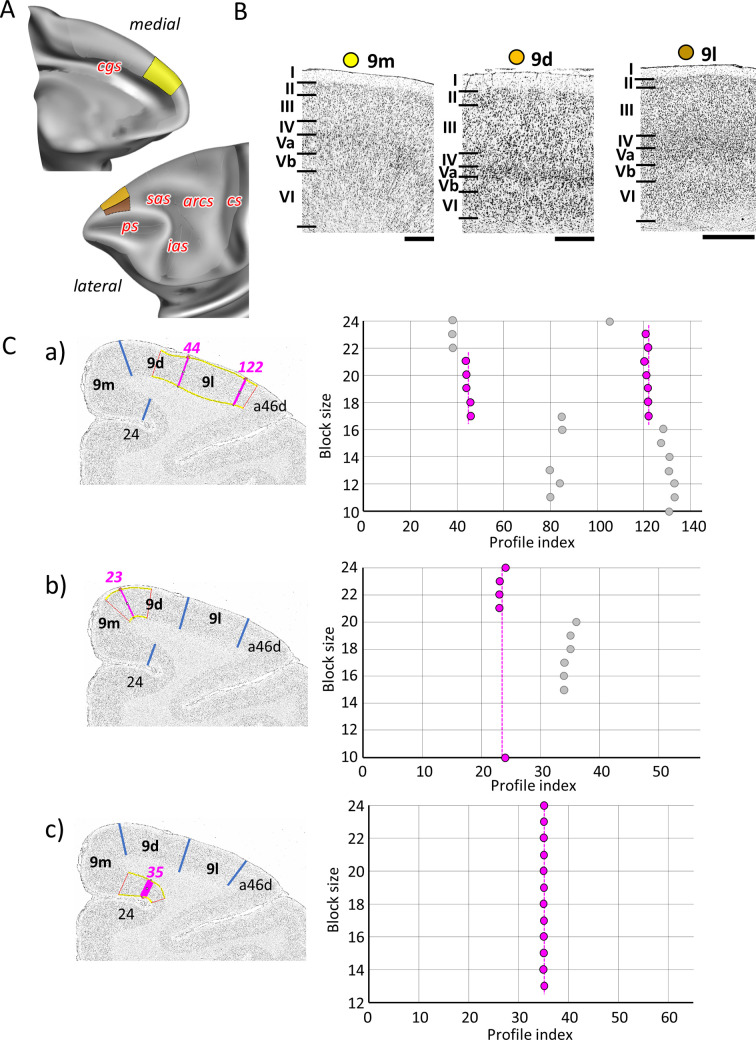
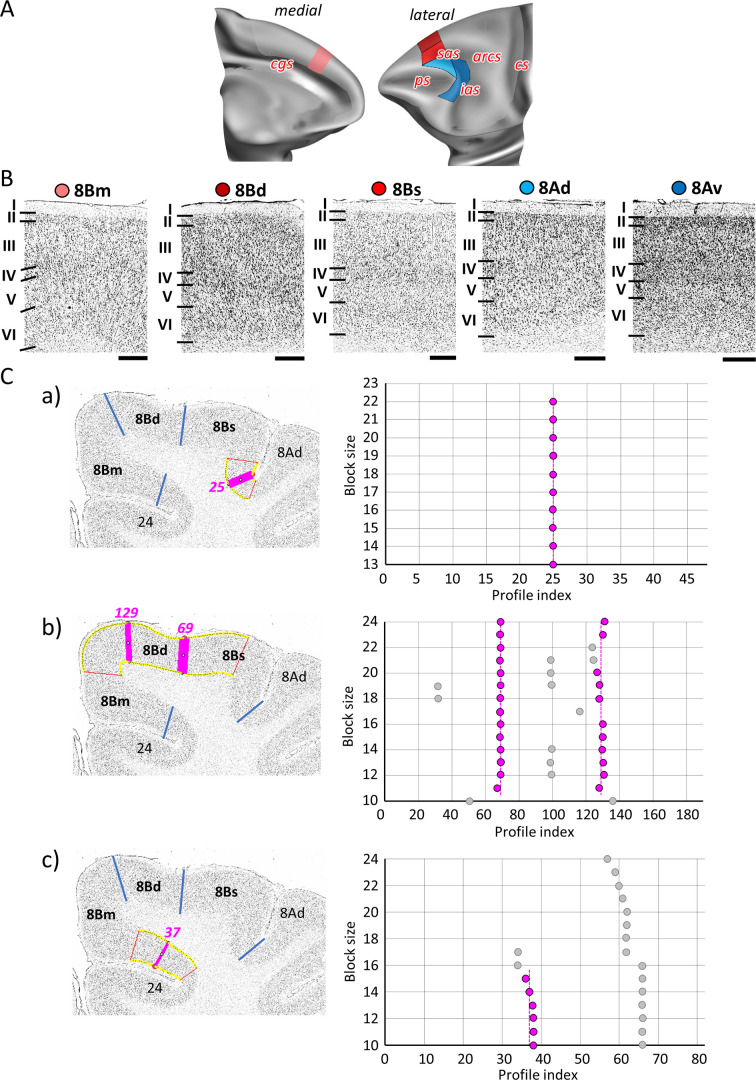
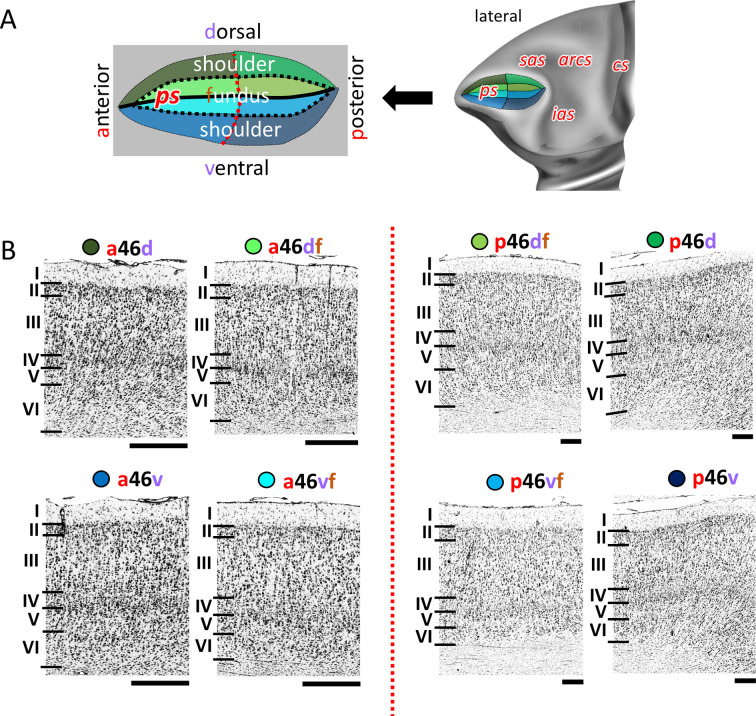
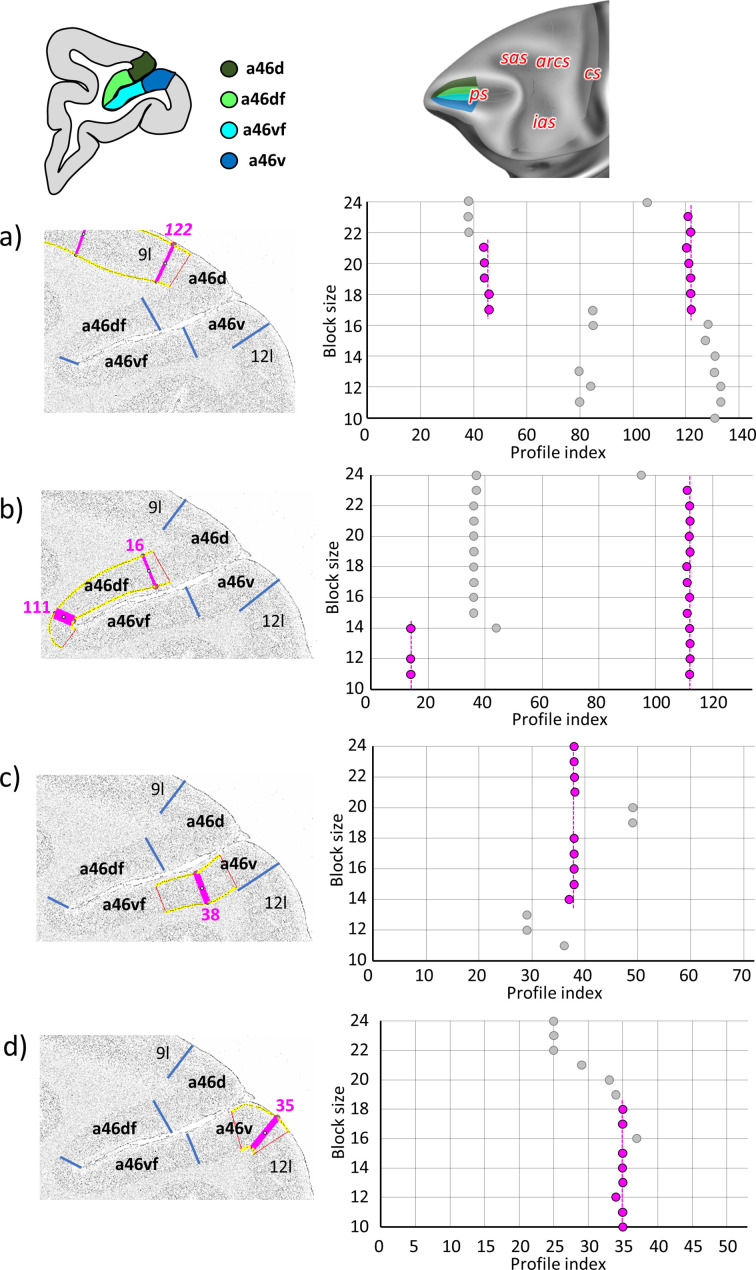
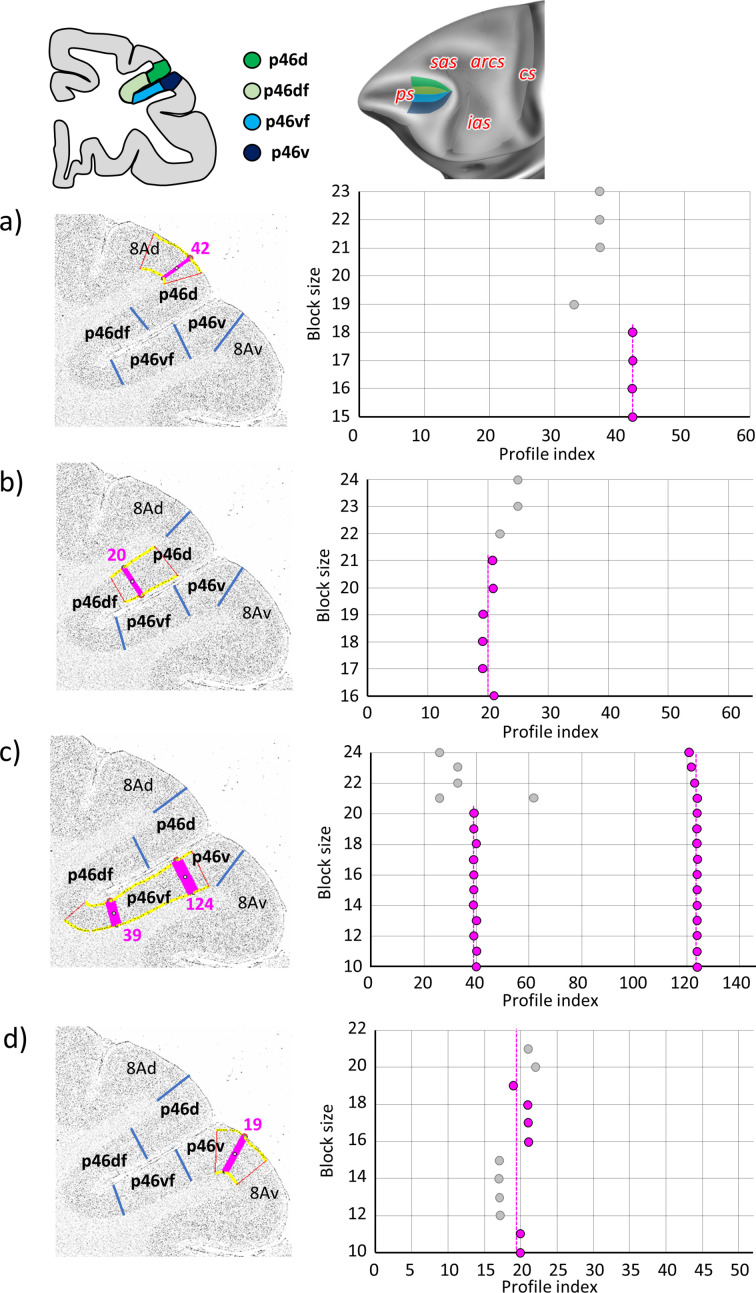
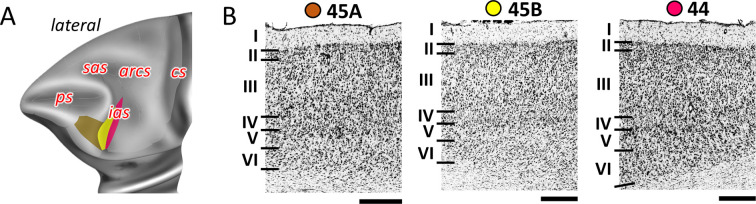
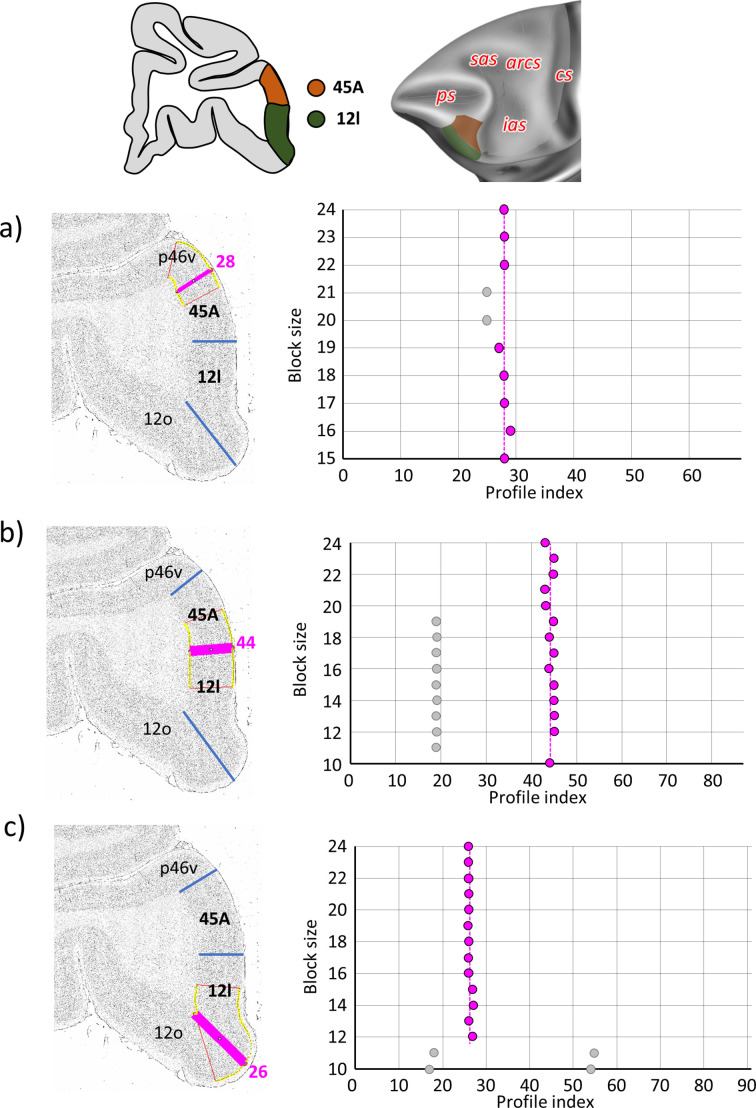
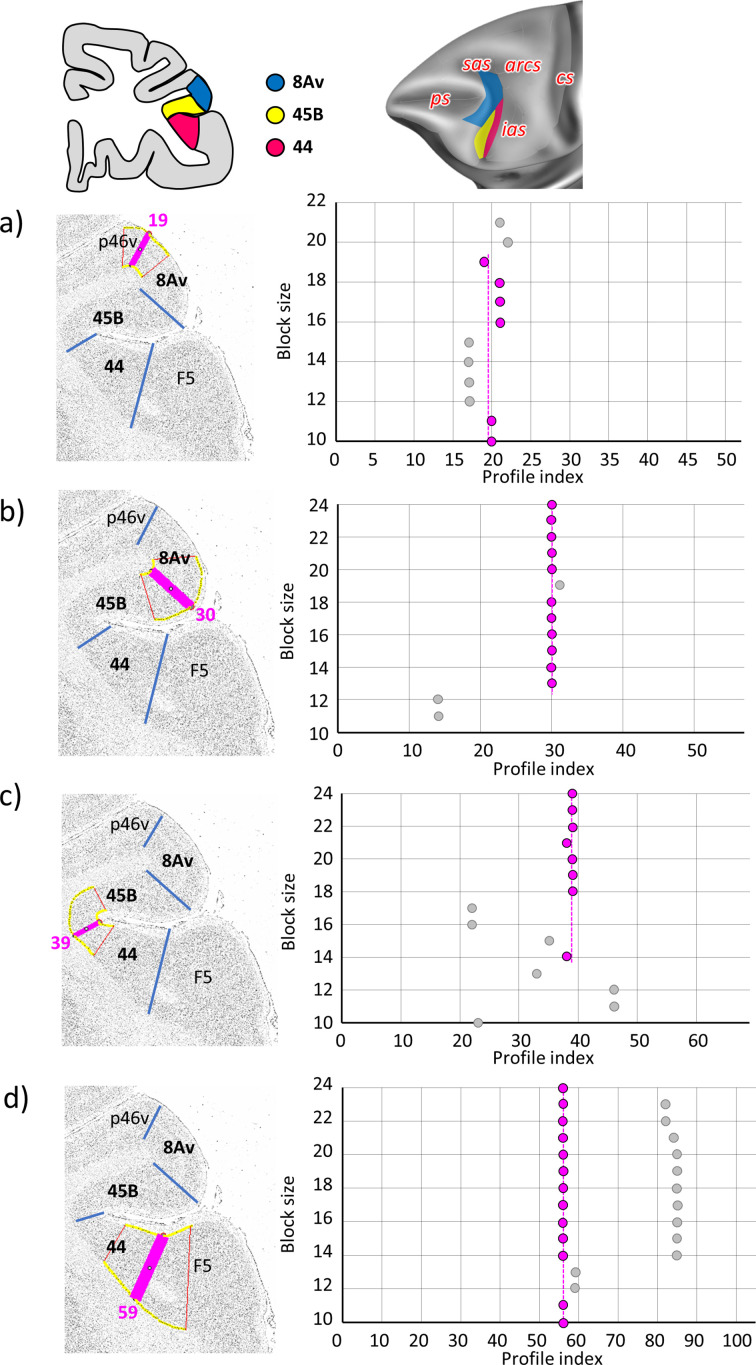
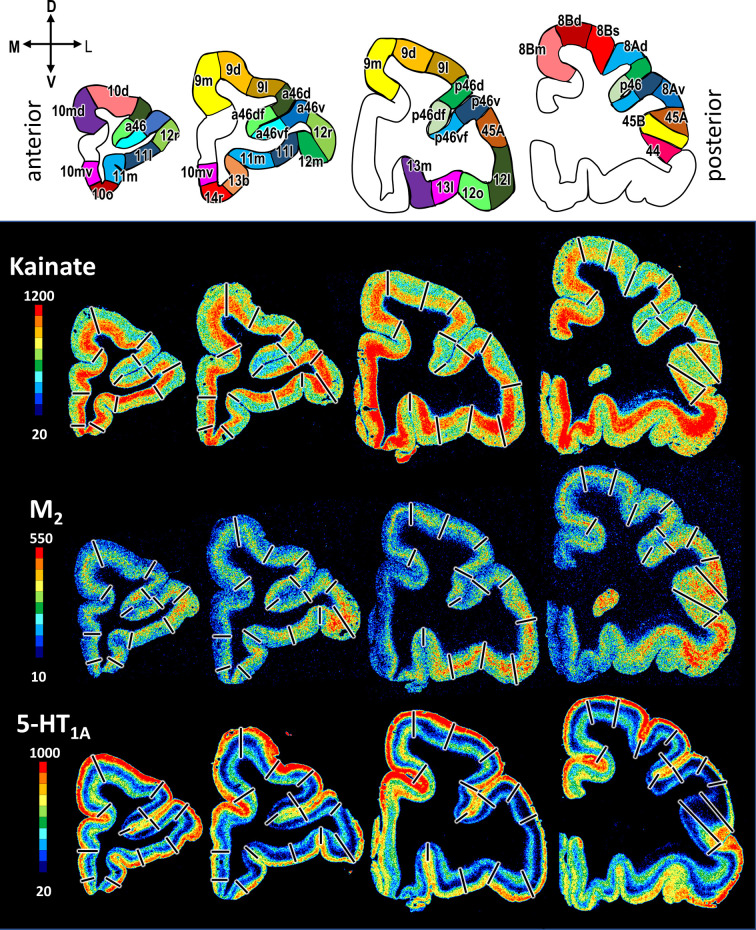
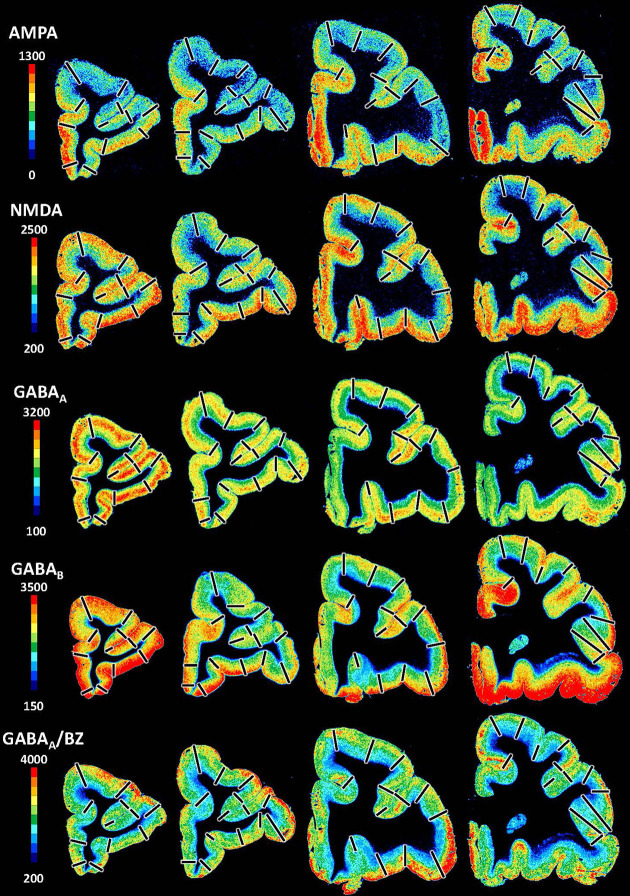
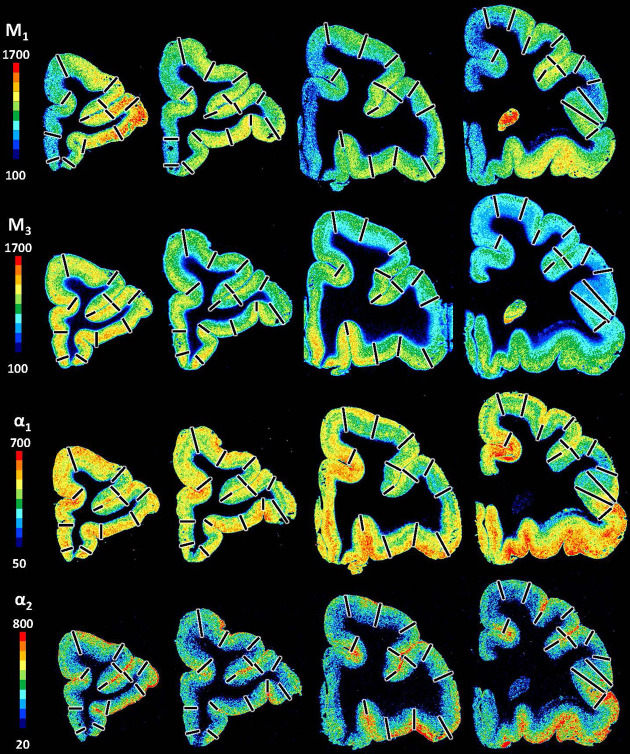
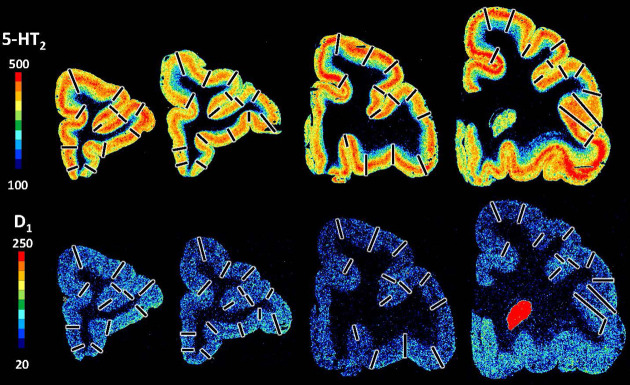
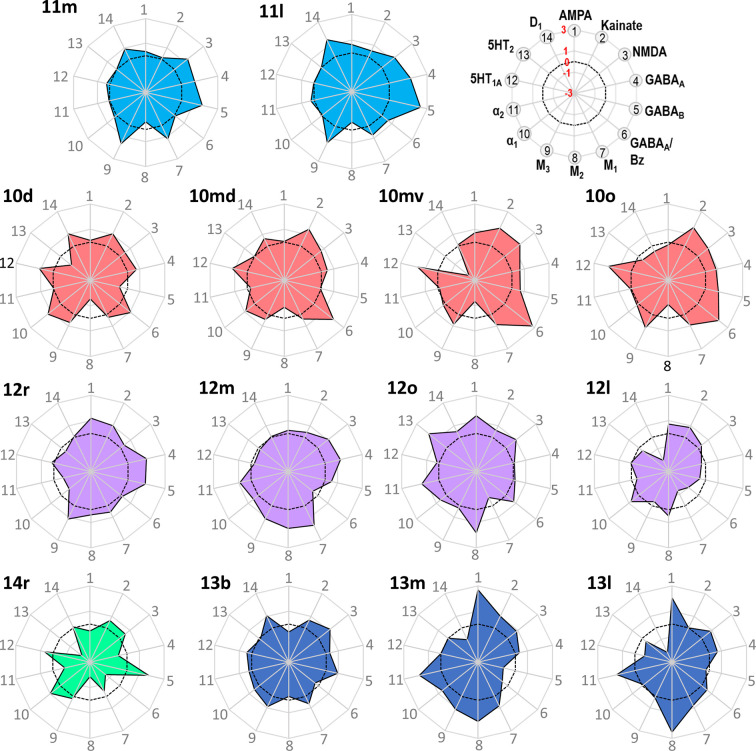
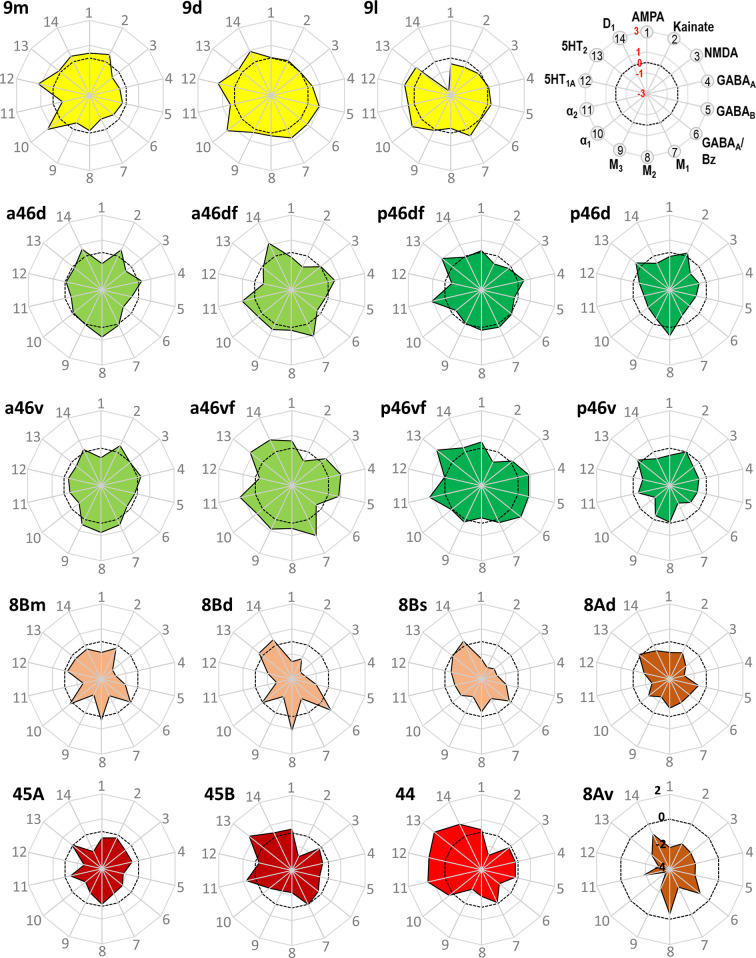
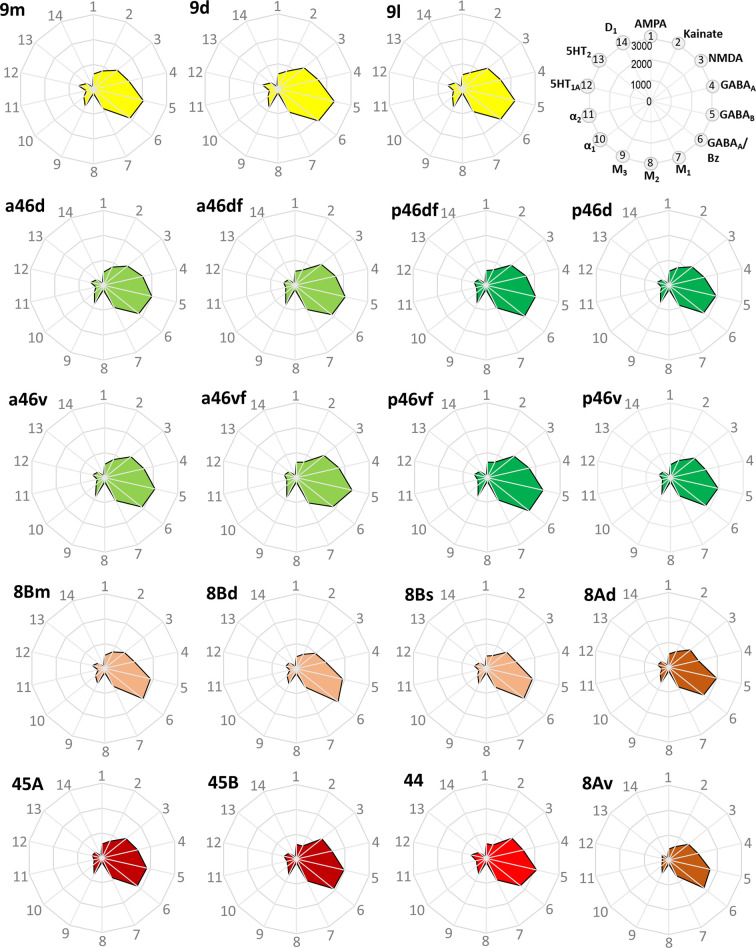
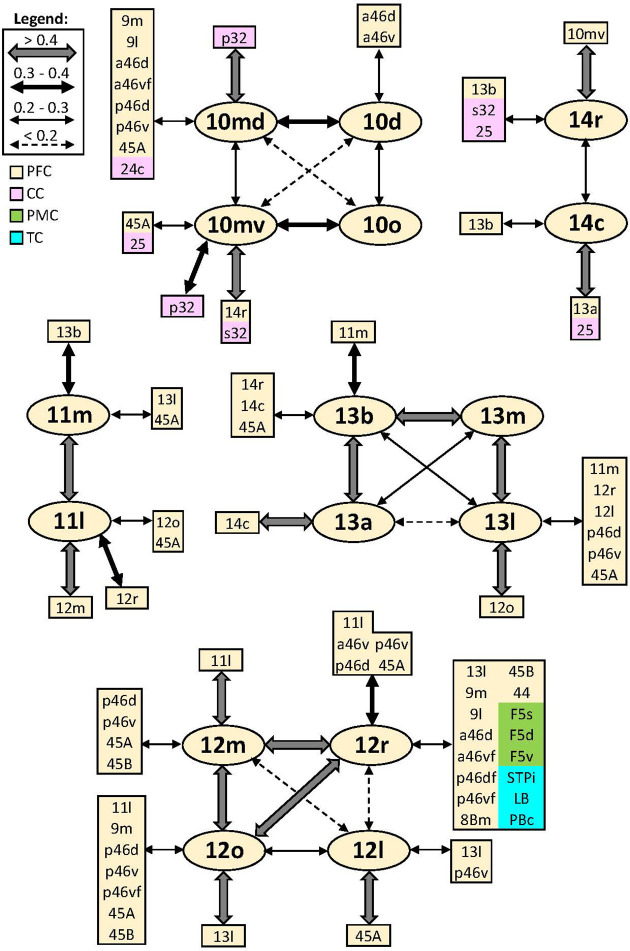
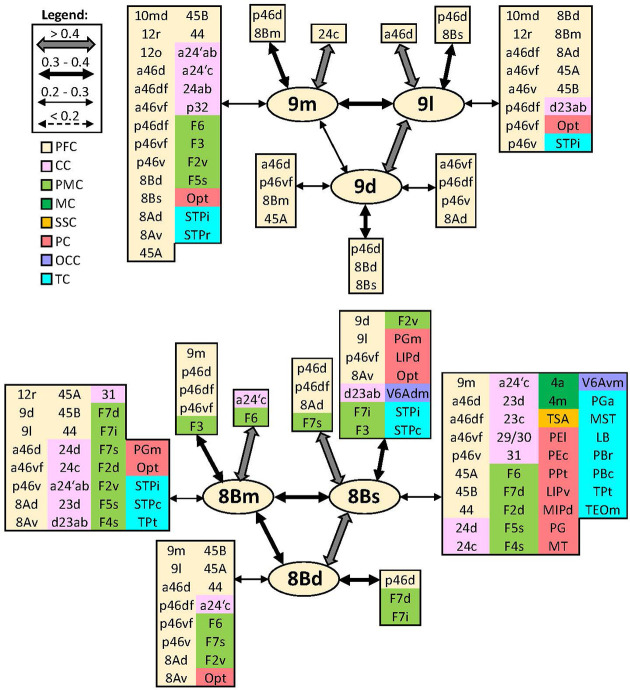
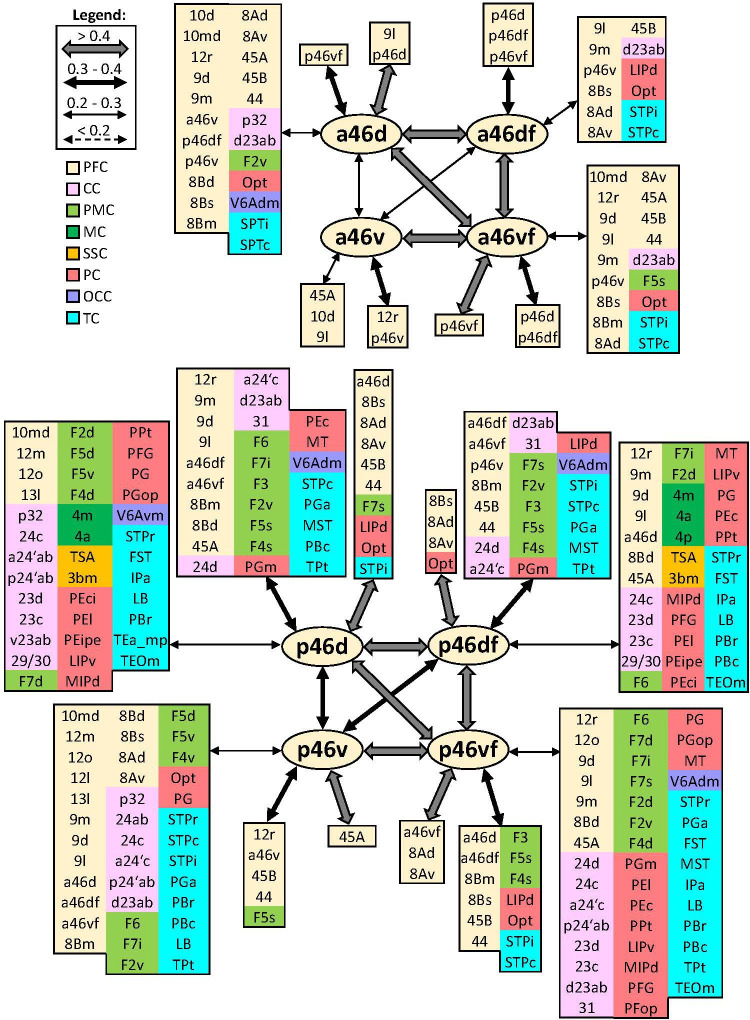
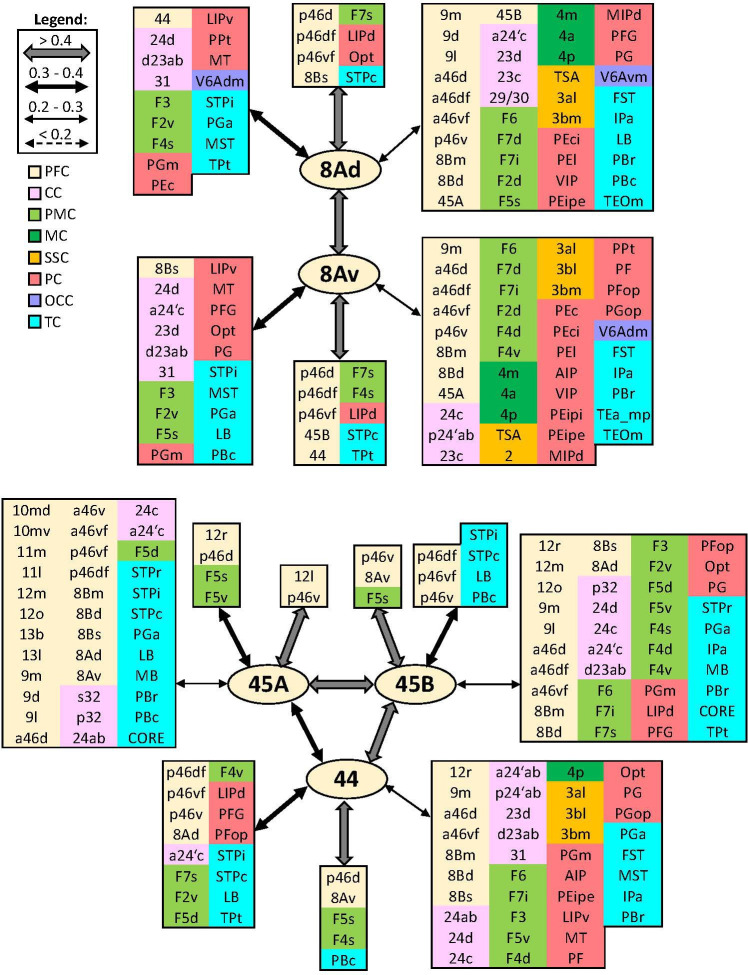
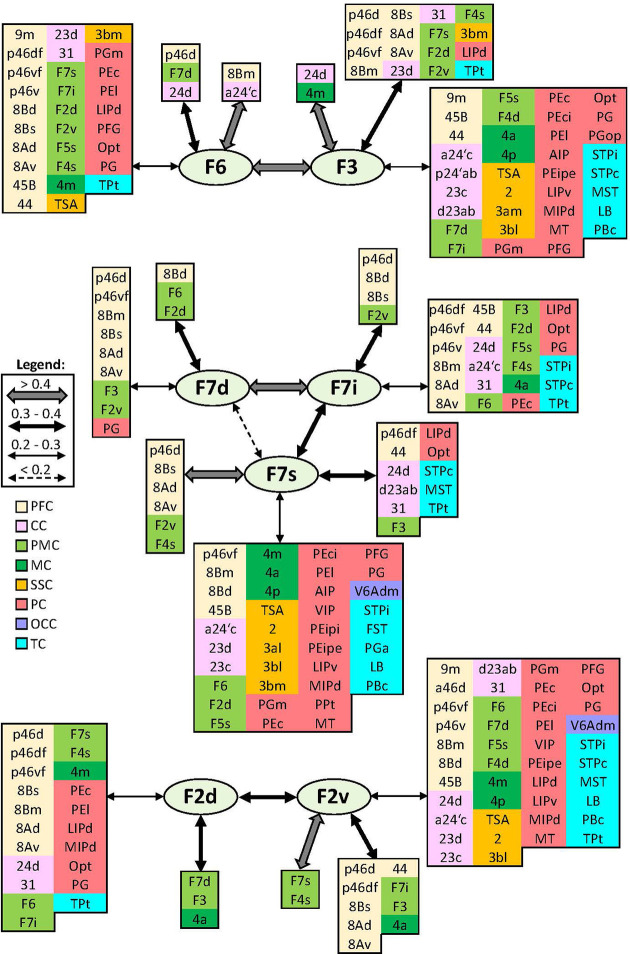
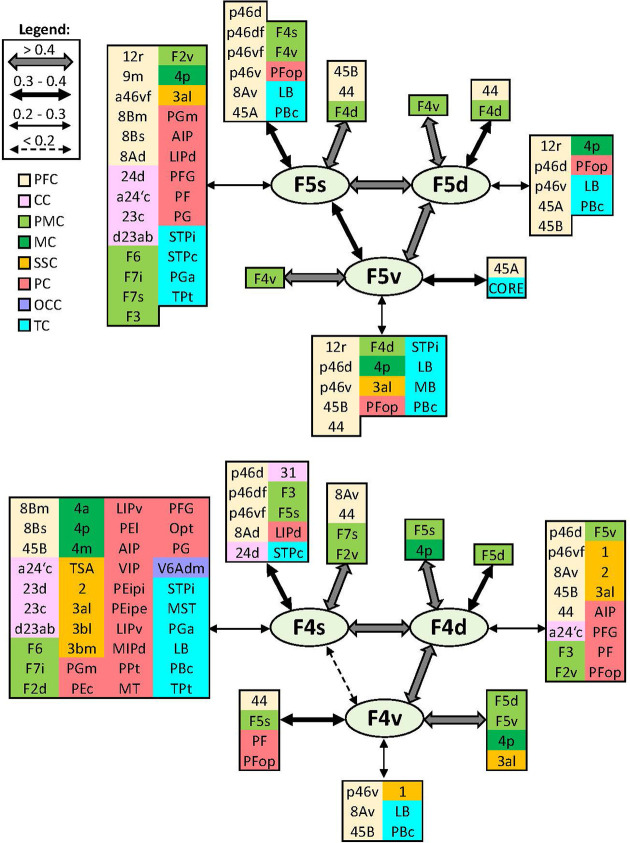
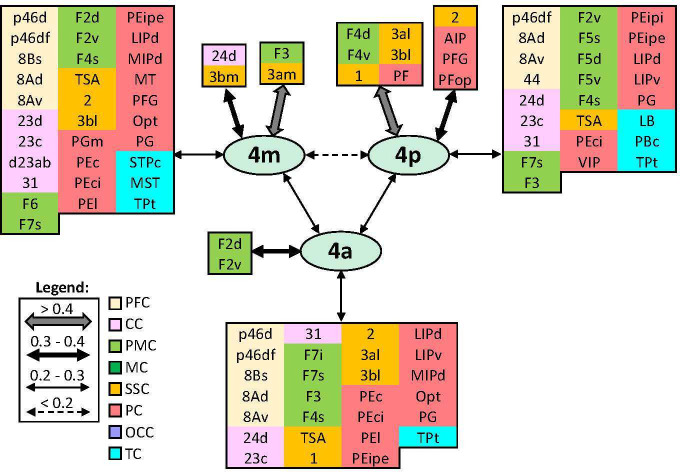
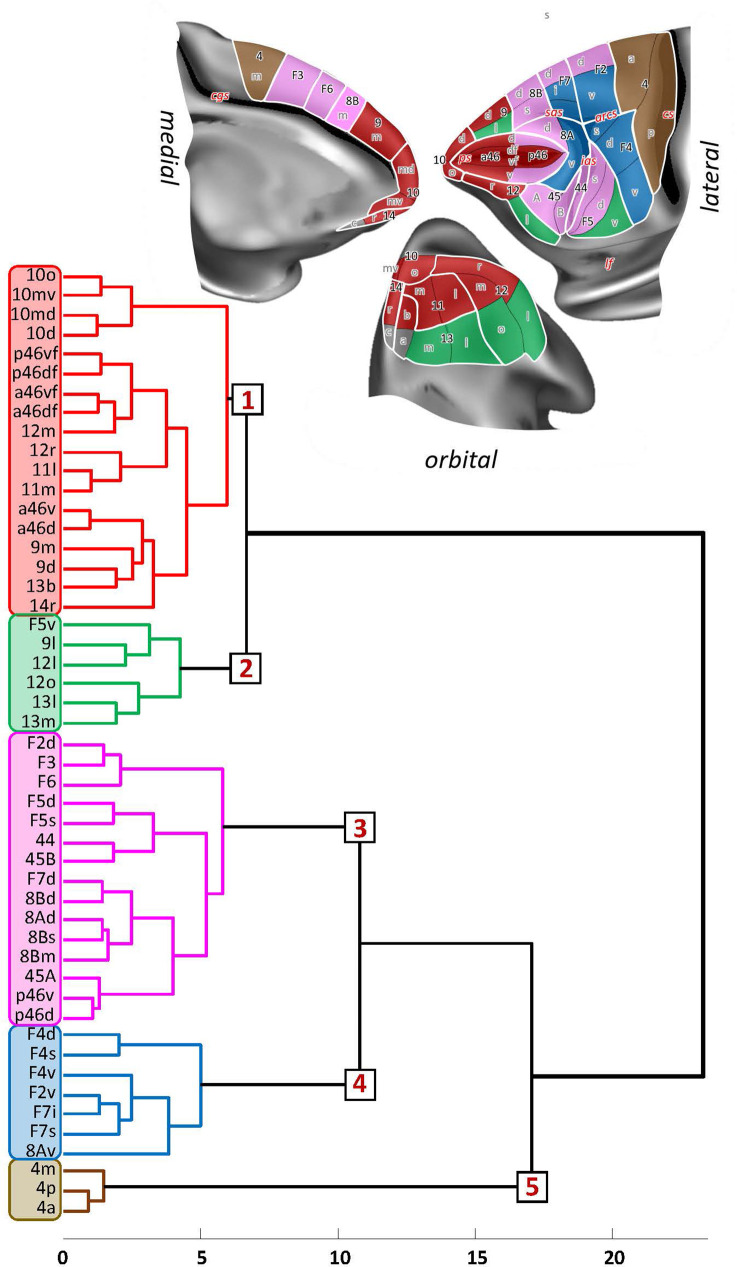
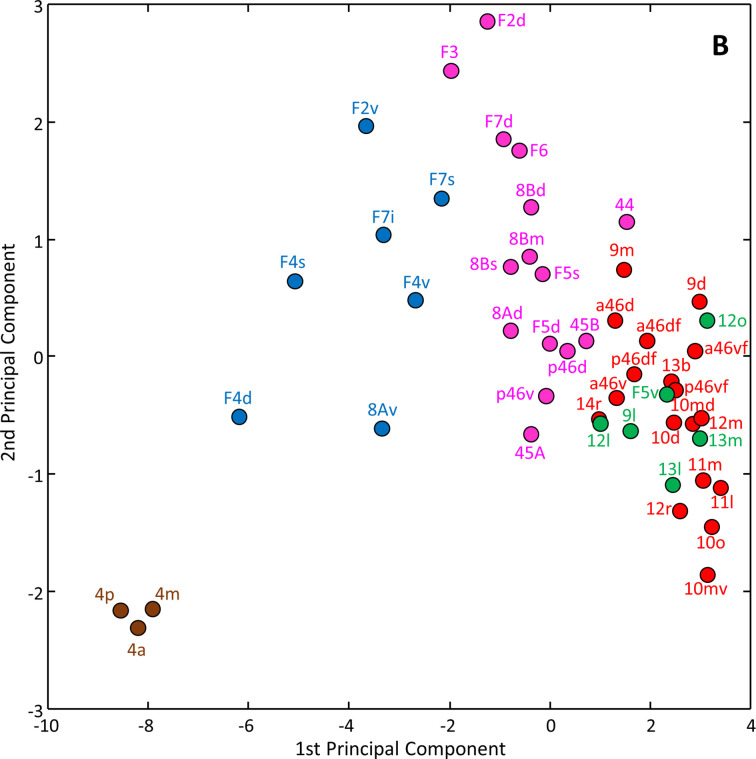
Based on quantitative cyto- and receptor architectonic analyses, we identified 35 prefrontal areas, including novel subdivisions of Walker's areas 10, 9, 8B, and 46. Statistical analysis of receptor densities revealed regional differences in lateral and ventrolateral prefrontal cortex. Indeed, structural and functional organization of subdivisions encompassing areas 46 and 12 demonstrated significant differences in the interareal levels of α receptors. Furthermore, multivariate analysis included receptor fingerprints of previously identified 16 motor areas in the same macaque brains and revealed 5 clusters encompassing frontal lobe areas. We used the MRI datasets from the non-human primate data sharing consortium PRIME-DE to perform functional connectivity analyses using the resulting frontal maps as seed regions. In general, rostrally located frontal areas were characterized by bigger fingerprints, that is, higher receptor densities, and stronger regional interconnections. Whereas more caudal areas had smaller fingerprints, but showed a widespread connectivity pattern with distant cortical regions. Taken together, this study provides a comprehensive insight into the molecular structure underlying the functional organization of the cortex and, thus, reconcile the discrepancies between the structural and functional hierarchical organization of the primate frontal lobe. Finally, our data are publicly available via the EBRAINS and BALSA repositories for the entire scientific community.
基于定量细胞和受体构筑分析,我们确定了 35 个前额叶区域,包括沃克氏区 10、9、8B 和 46 的新细分区域。受体密度的统计分析显示外侧和腹外侧前额叶皮层存在区域差异。事实上,包括区域 46 和 12 在内的细分区域的结构和功能组织在α受体的区域间水平上表现出显著差异。此外,多元分析包括同一猕猴大脑中先前确定的 16 个运动区域的受体指纹,并揭示了包含额叶区域的 5 个簇。我们使用非人类灵长类动物数据共享联盟 PRIME-DE 的 MRI 数据集,使用生成的额叶图谱作为种子区域进行功能连接分析。一般来说,位于前部的区域具有更大的指纹,即更高的受体密度和更强的区域内连接。而更靠后的区域具有较小的指纹,但与远处的皮质区域显示出广泛的连接模式。总的来说,这项研究提供了对皮层功能组织背后的分子结构的全面了解,从而调和了灵长类动物额叶的结构和功能层次组织之间的差异。最后,我们的数据通过 EBRAINS 和 BALSA 存储库向整个科学界公开。