Konno K, Morales M F
Proc Natl Acad Sci U S A. 1985 Dec;82(23):7904-8. doi: 10.1073/pnas.82.23.7904.
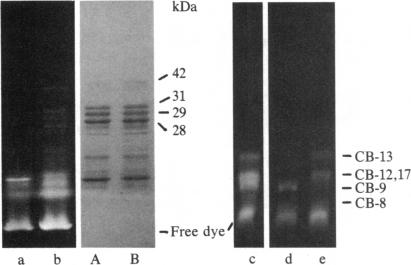
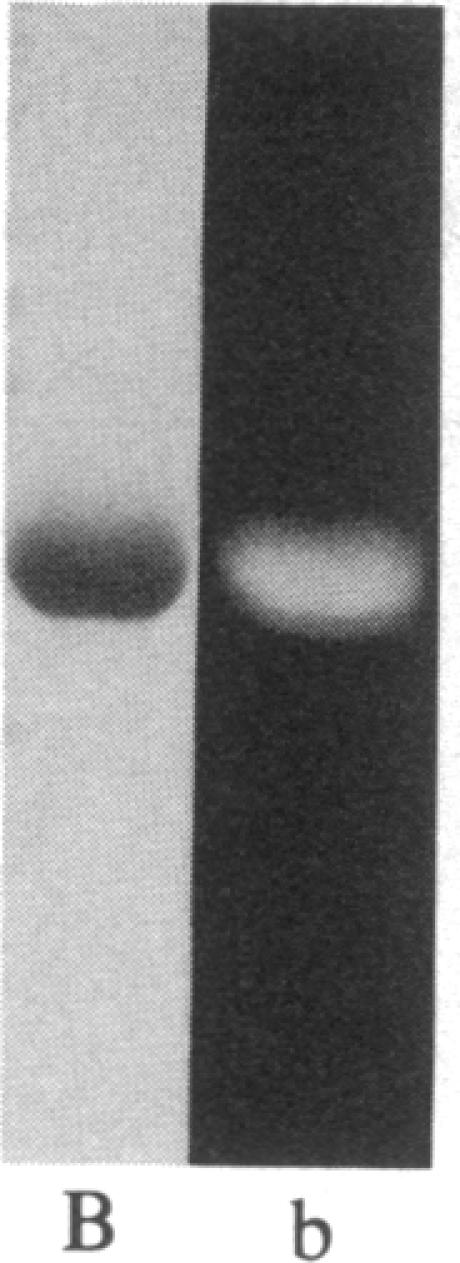
The removal of bound metal ions from G-actin uncovered two thiols, Cys-10 and Cys-257. The uncovering of these thiols requires a free calcium concentration lower than 10 nM. Therefore, participation of one or both thiols in Ca2+ binding is suggested. Actin labeled with N-(5-fluoresceinyl)maleimide in the absence of calcium moves as a doublet in NaDodSO4/PAGE. It is suggested that two conformers are induced by metal removal and labeling.
从G-肌动蛋白中去除结合的金属离子后发现了两个巯基,即半胱氨酸-10和半胱氨酸-257。这些巯基的暴露需要游离钙浓度低于10 nM。因此,提示一个或两个巯基参与了Ca2+结合。在没有钙的情况下用N-(5-荧光素基)马来酰亚胺标记的肌动蛋白在NaDodSO4/PAGE中以双峰形式移动。提示金属去除和标记诱导了两种构象体。