Department of Visceral Surgery and Medicine, Inselspital, Bern University Hospital, University Hospital of Bern, Bern, Switzerland.
Department for BioMedical Research, Visceral Surgery and Medicine, University Hospital of Bern, Bern, Switzerland.
Elife. 2024 Aug 20;13:RP96678. doi: 10.7554/eLife.96678.
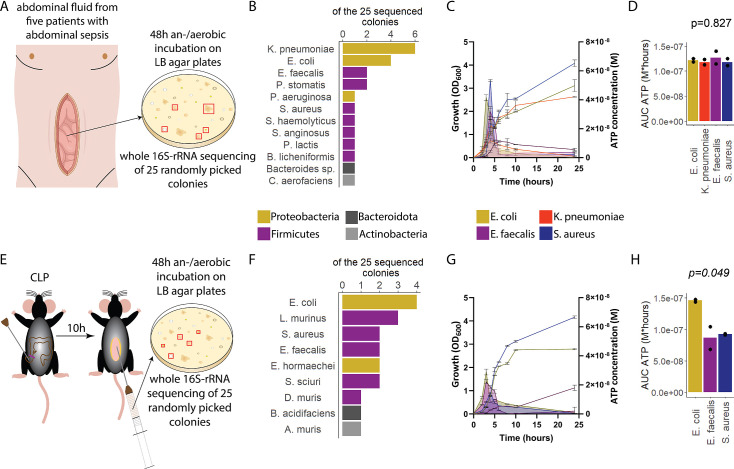
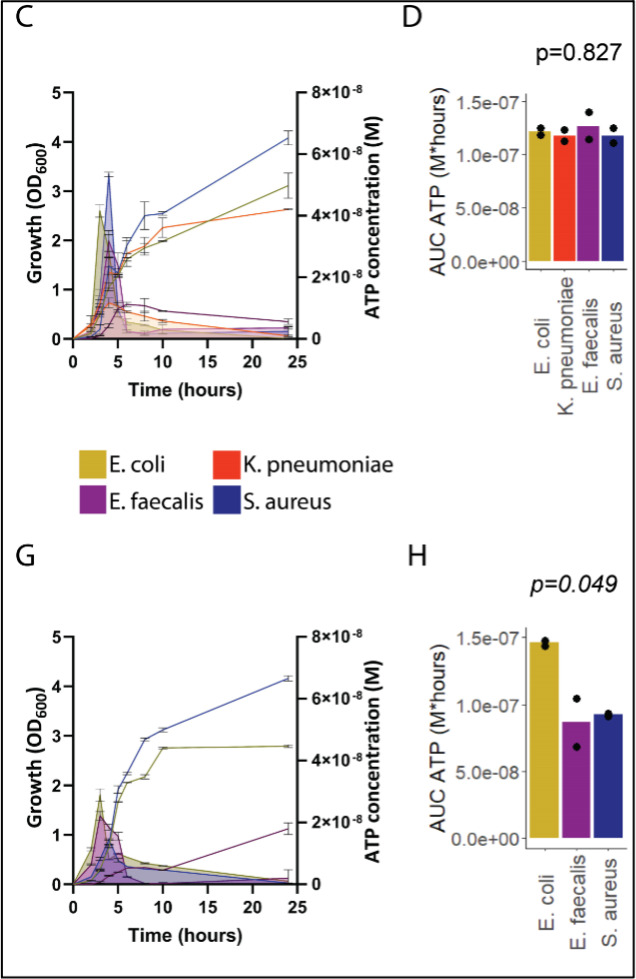
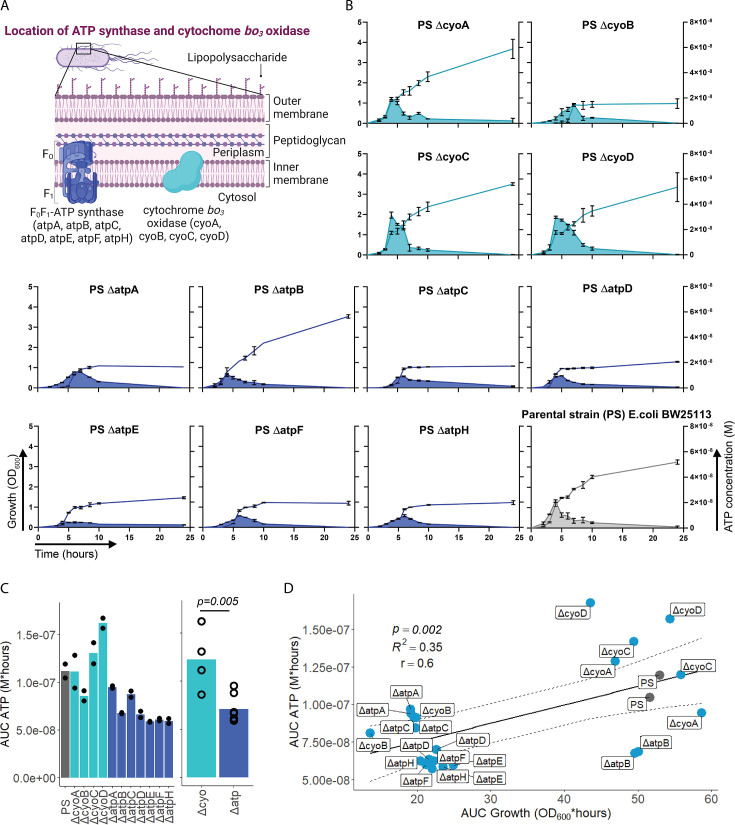
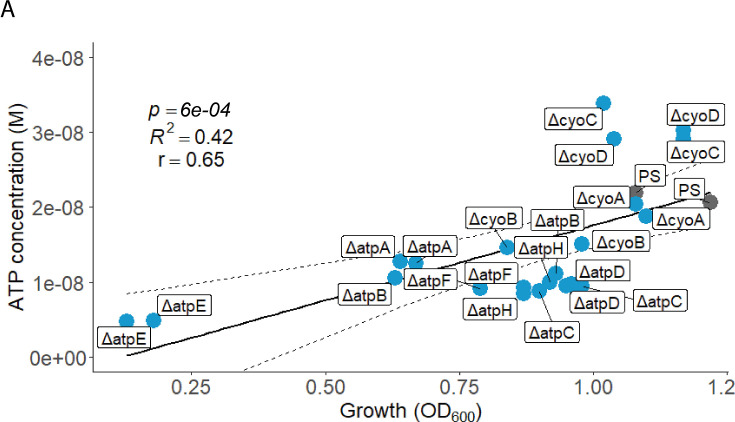
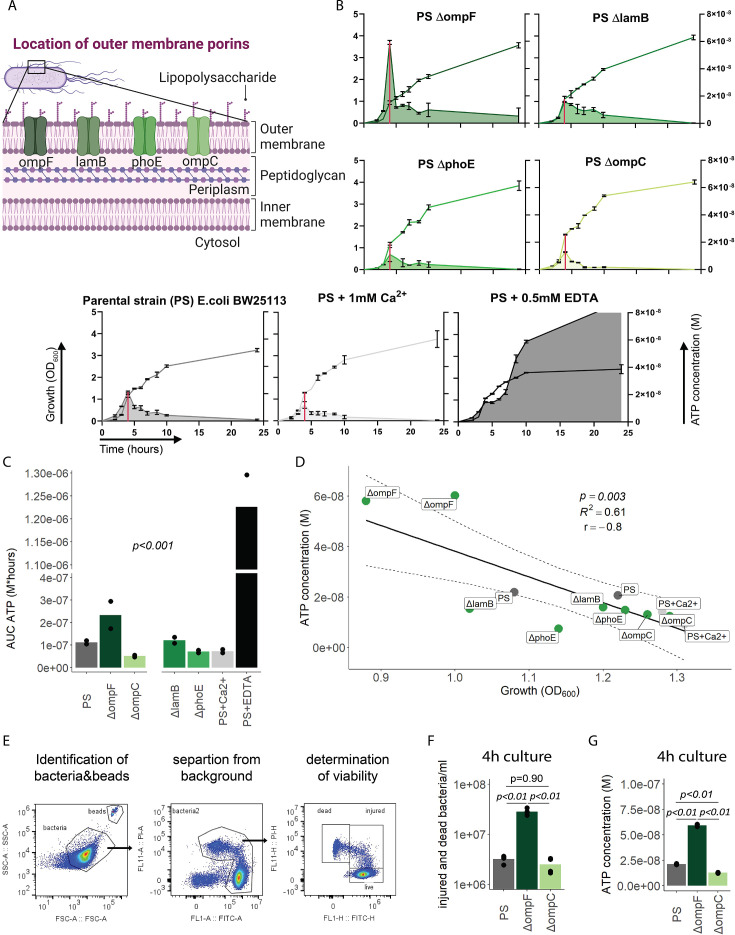
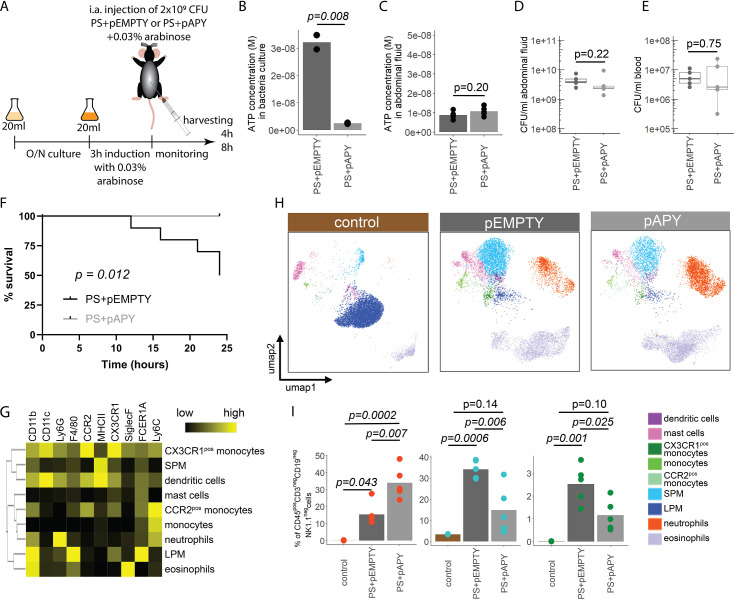
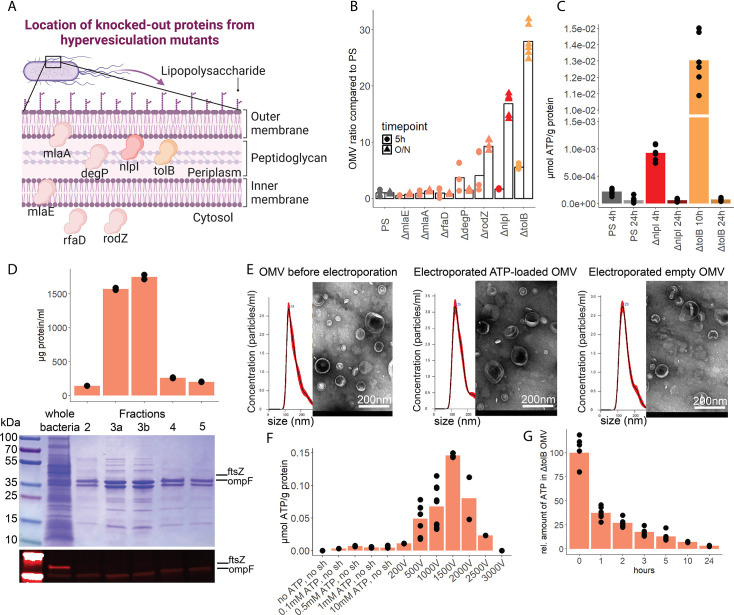
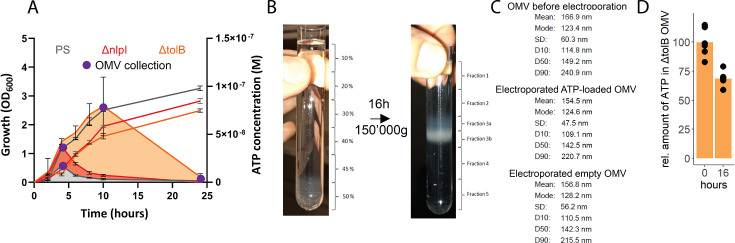
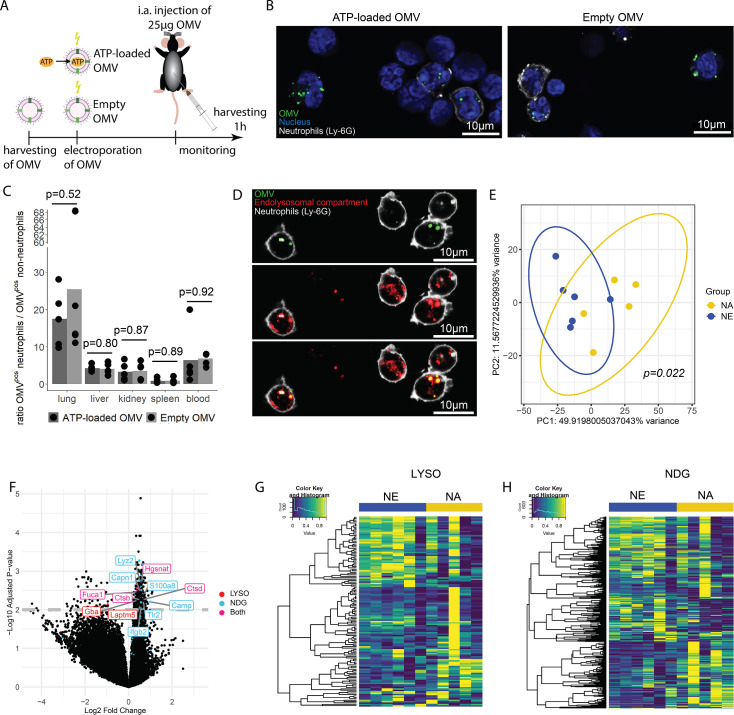
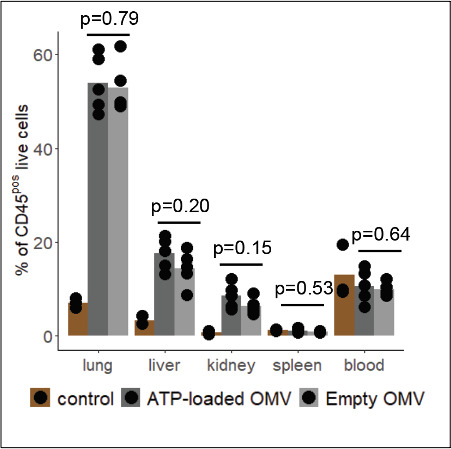
Sepsis causes millions of deaths per year worldwide and is a current global health priority declared by the WHO. Sepsis-related deaths are a result of dysregulated inflammatory immune responses indicating the need to develop strategies to target inflammation. An important mediator of inflammation is extracellular adenosine triphosphate (ATP) that is released by inflamed host cells and tissues, and also by bacteria in a strain-specific and growth-dependent manner. Here, we investigated the mechanisms by which bacteria release ATP. Using genetic mutant strains of (), we demonstrate that ATP release is dependent on ATP synthase within the inner bacterial membrane. In addition, impaired integrity of the outer bacterial membrane notably contributes to ATP release and is associated with bacterial death. In a mouse model of abdominal sepsis, local effects of bacterial ATP were analyzed using a transformed bearing an arabinose-inducible periplasmic apyrase hydrolyzing ATP to be released. Abrogating bacterial ATP release shows that bacterial ATP suppresses local immune responses, resulting in reduced neutrophil counts and impaired survival. In addition, bacterial ATP has systemic effects via its transport in outer membrane vesicles (OMV). ATP-loaded OMV are quickly distributed throughout the body and upregulated expression of genes activating degranulation in neutrophils, potentially contributing to the exacerbation of sepsis severity. This study reveals mechanisms of bacterial ATP release and its local and systemic roles in sepsis pathogenesis.
败血症每年导致全球数百万人死亡,是世卫组织宣布的当前全球卫生重点。与败血症相关的死亡是由于炎症免疫反应失调所致,这表明需要制定针对炎症的策略。炎症的一个重要介质是细胞外三磷酸腺苷 (ATP),它由炎症宿主细胞和组织释放,细菌也以菌株特异性和生长依赖性的方式释放。在这里,我们研究了细菌释放 ATP 的机制。使用 ( ) 的遗传突变株,我们证明 ATP 的释放依赖于内膜中的 ATP 合酶。此外,细菌外膜完整性的受损显著促进了 ATP 的释放,并与细菌死亡相关。在腹部败血症的小鼠模型中,使用携带阿拉伯糖诱导的周质质膜内的外切核酸酶水解释放的 ATP 的转化 来分析细菌 ATP 的局部作用。消除细菌 ATP 的释放表明细菌 ATP 抑制了局部免疫反应,导致中性粒细胞计数减少和存活率降低。此外,细菌 ATP 通过其在外膜囊泡 (OMV) 中的运输具有全身作用。负载 ATP 的 OMV 迅速分布在全身,并上调中性粒细胞脱颗粒的基因表达,可能导致败血症严重程度恶化。这项研究揭示了细菌 ATP 释放的机制及其在败血症发病机制中的局部和全身作用。