Department of Dermatology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA.
Center for Cellular Immunotherapies, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA.
Nature. 2024 Sep;633(8030):670-677. doi: 10.1038/s41586-024-07862-7. Epub 2024 Aug 28.
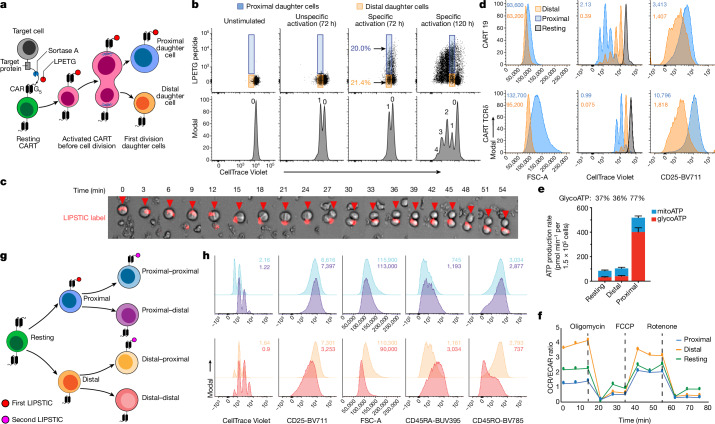
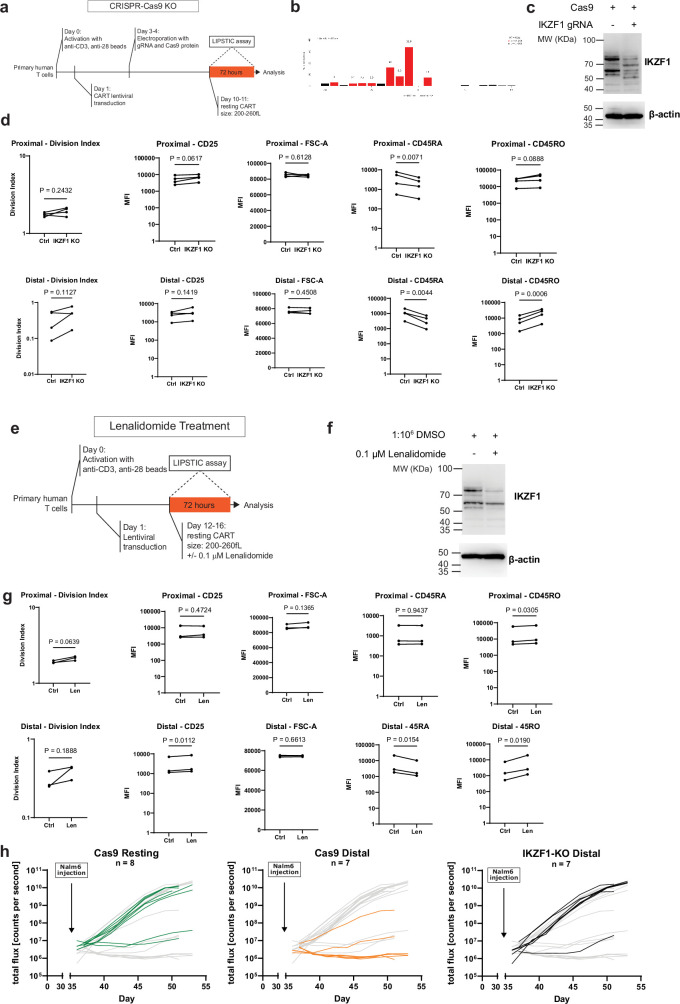
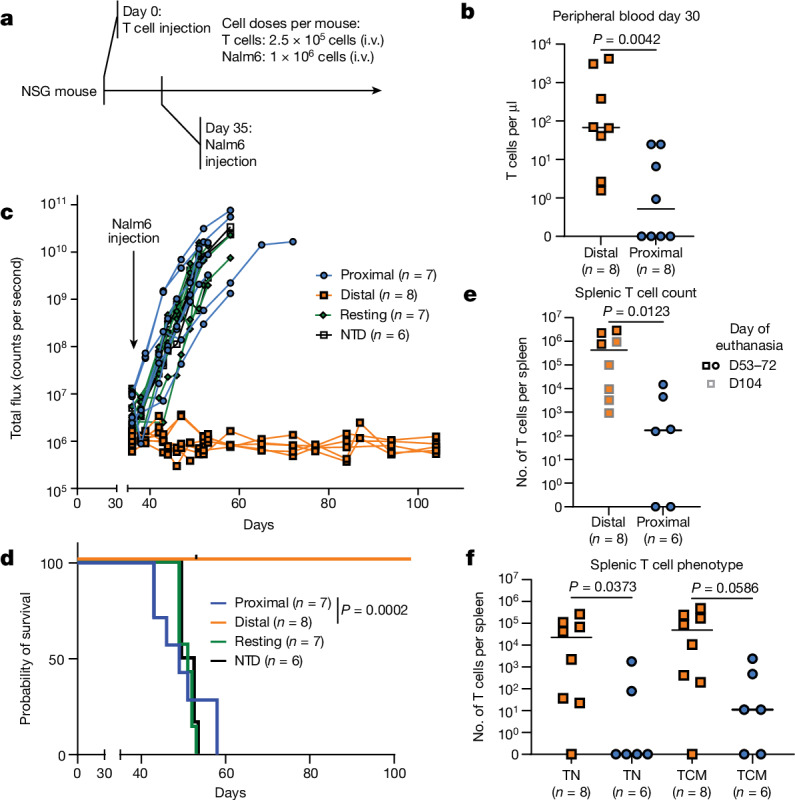
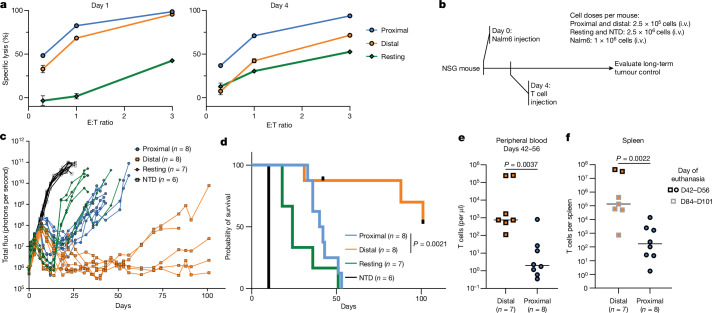
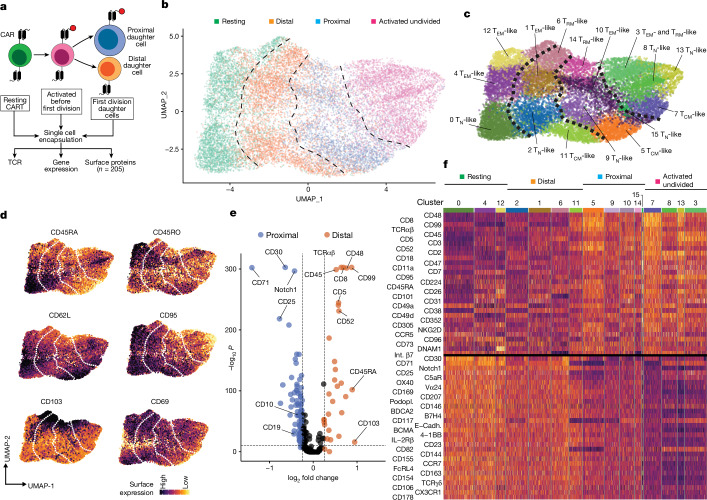
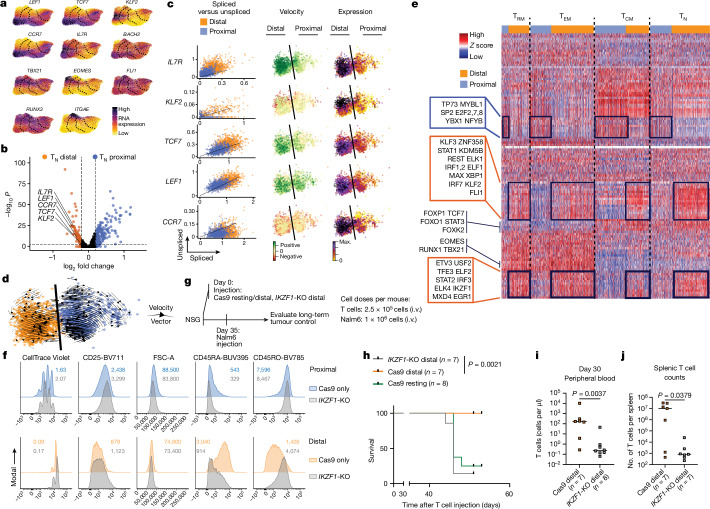
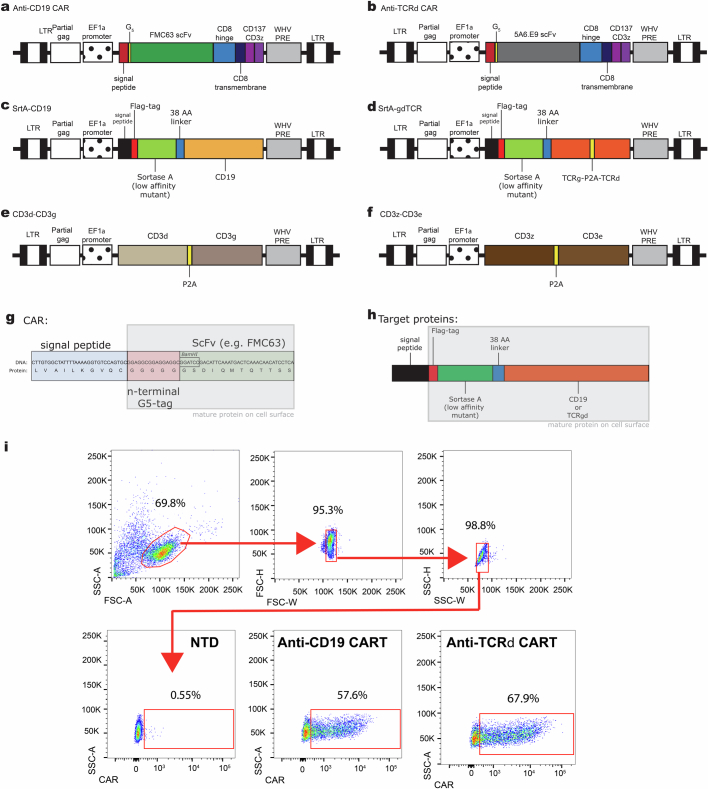
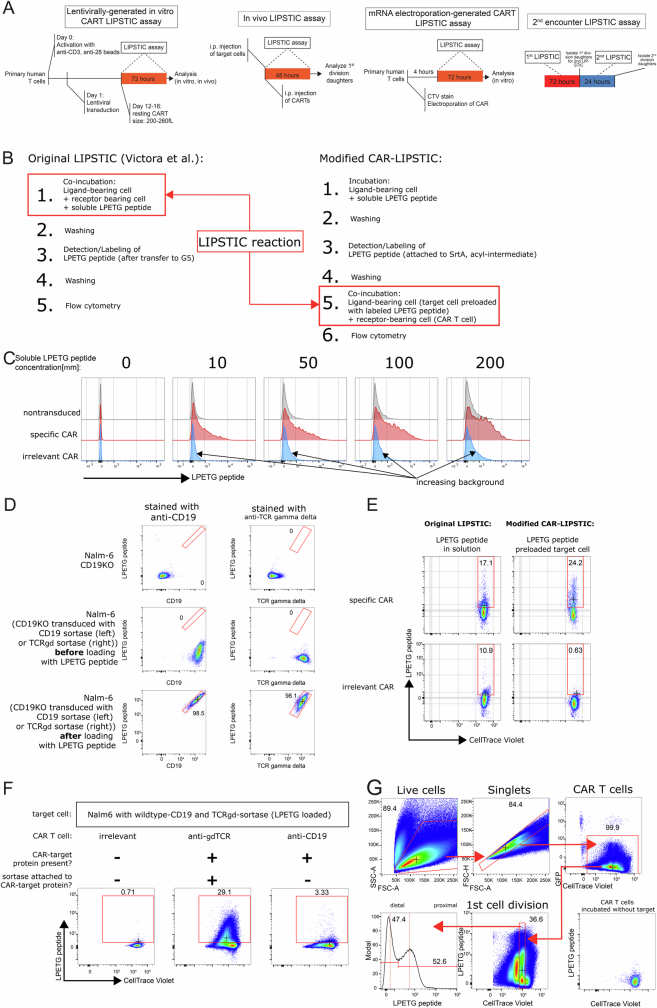
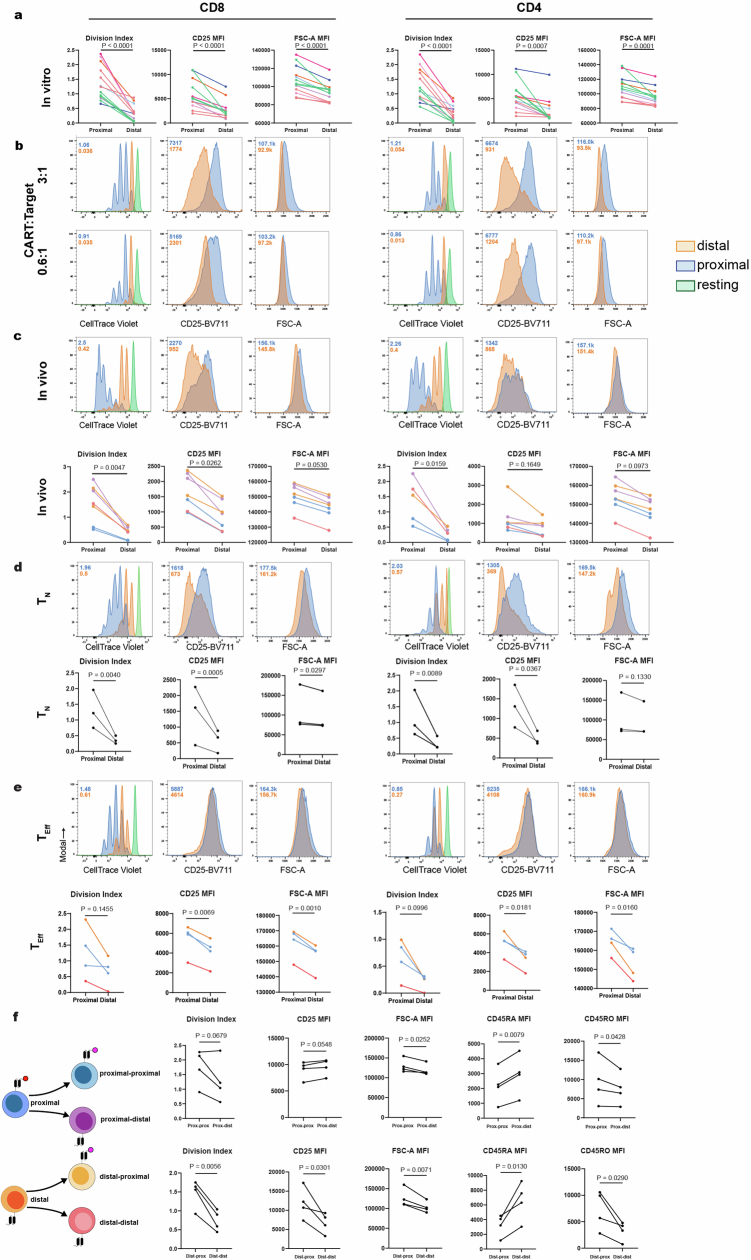
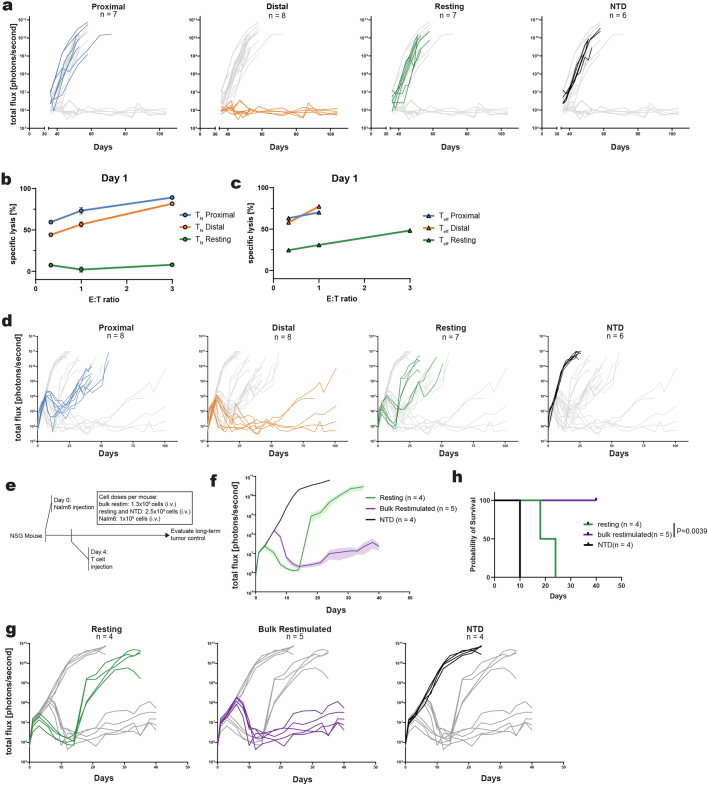
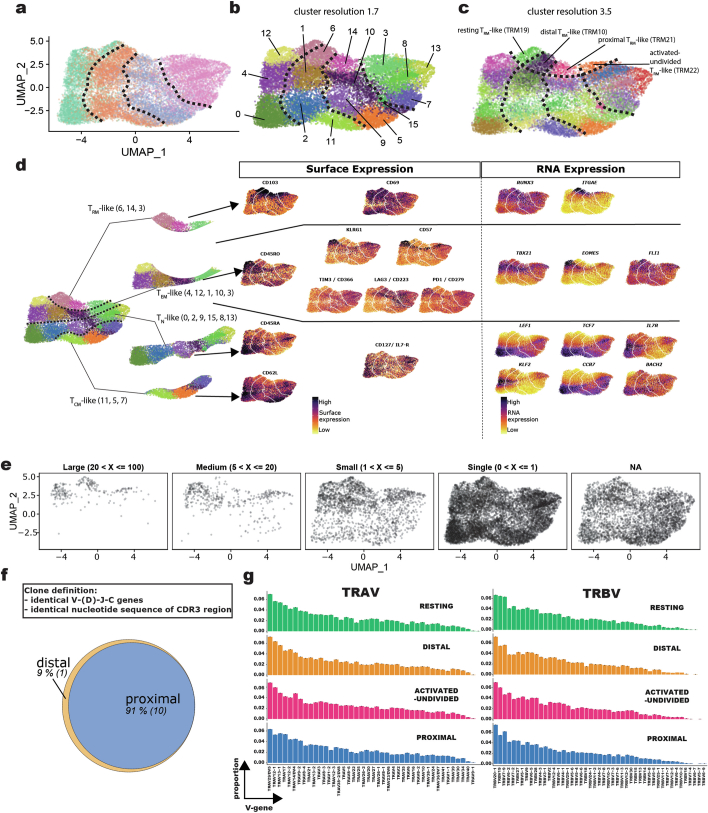
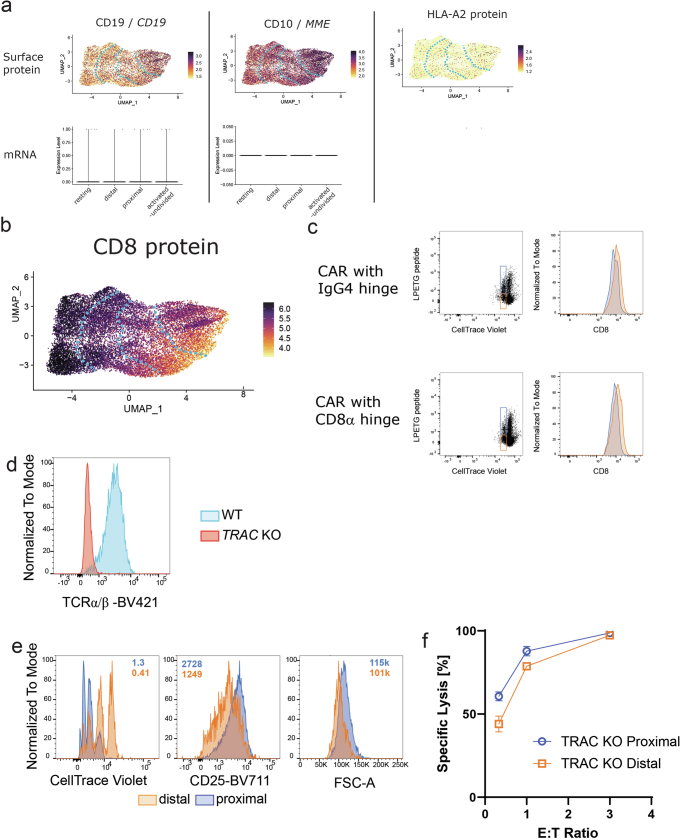
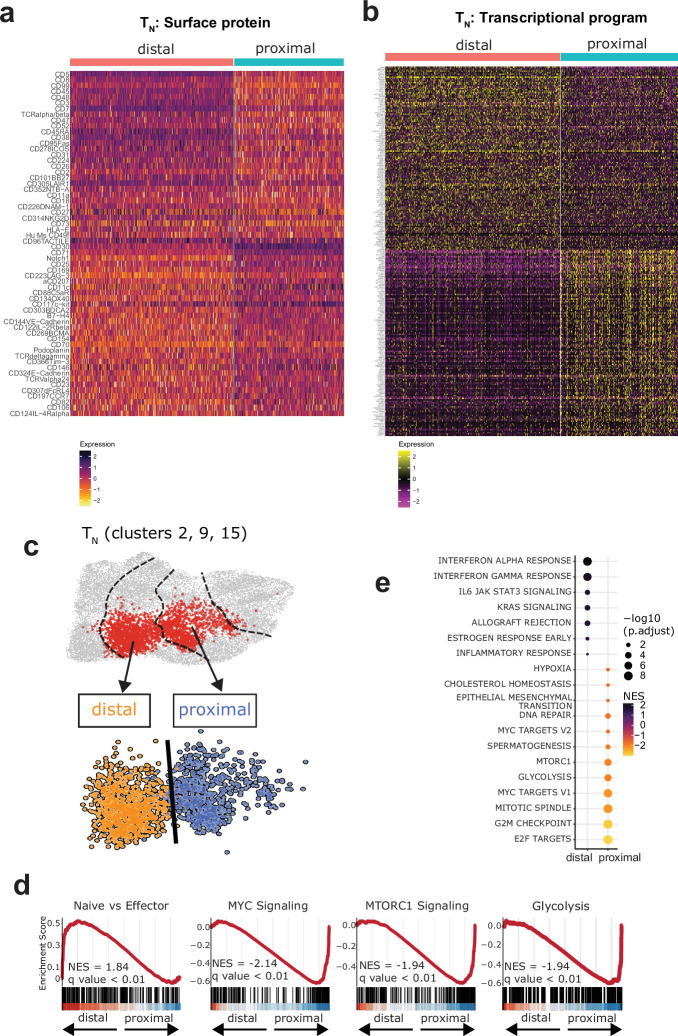
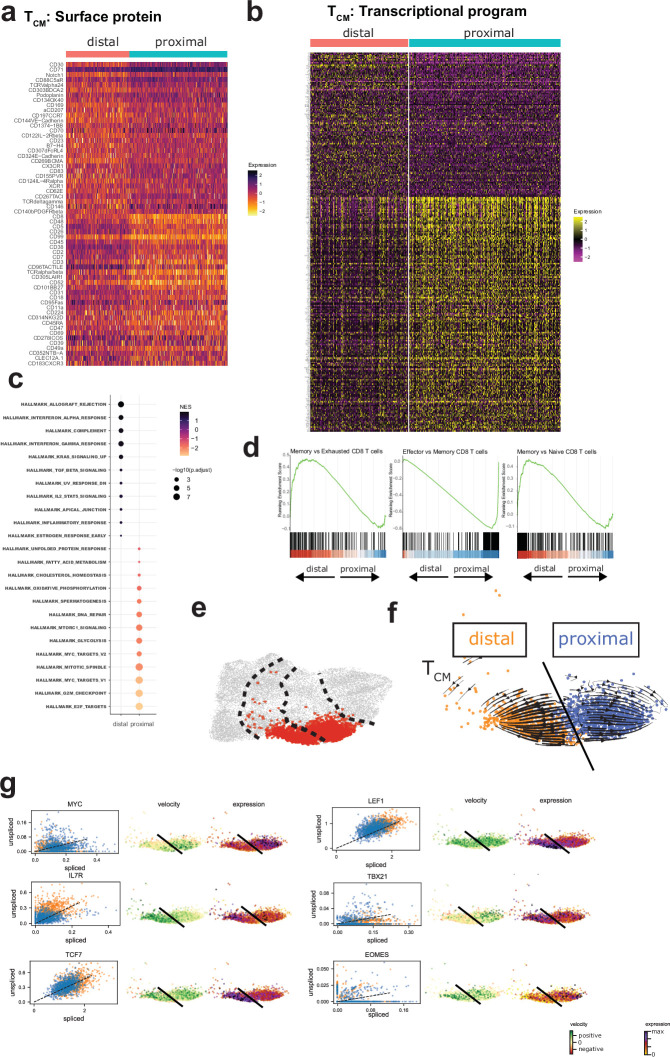
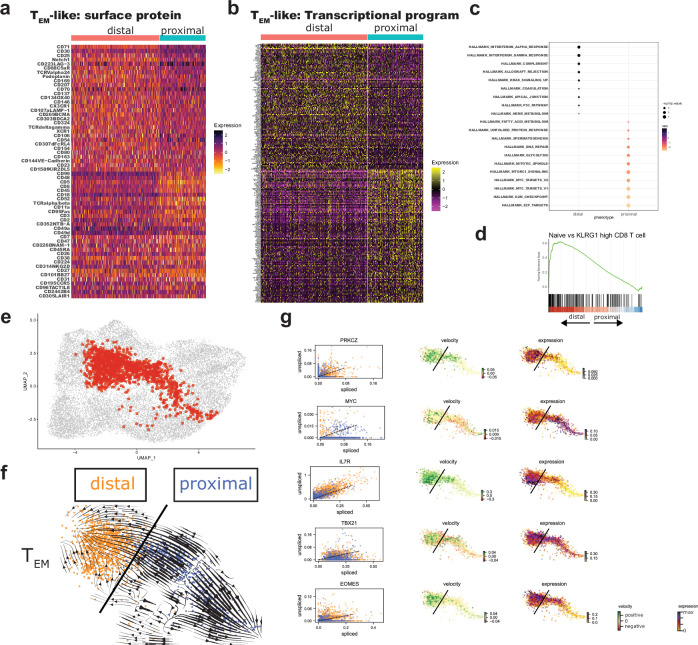
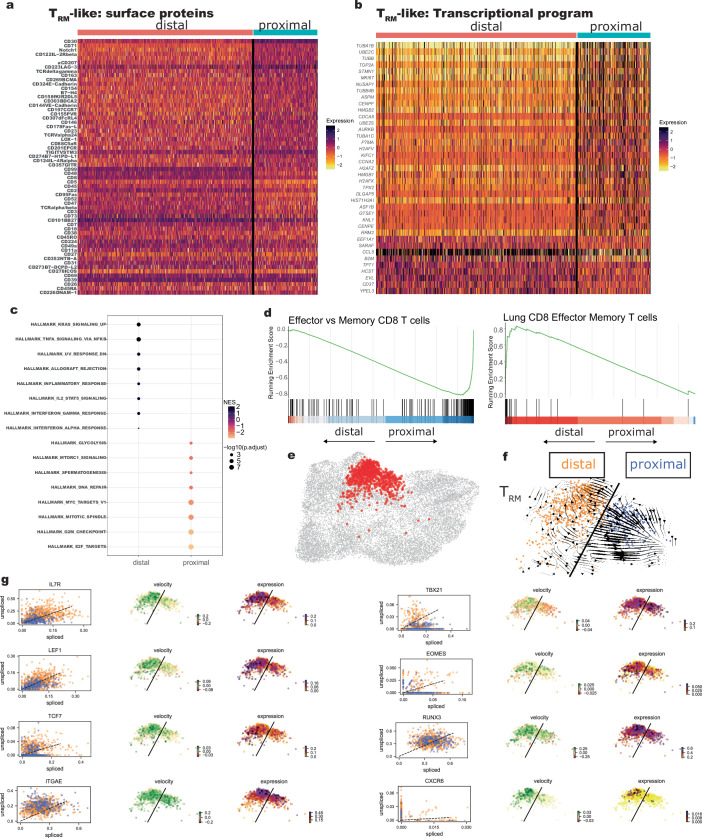
Early expansion and long-term persistence predict efficacy of chimeric antigen receptor T cells (CARTs), but mechanisms governing effector versus memory CART differentiation and whether asymmetric cell division induces differential fates in human CARTs remain unclear. Here we show that target-induced proximity labelling enables isolation of first-division proximal-daughter and distal-daughter CD8 CARTs that asymmetrically distribute their surface proteome and transcriptome, resulting in divergent fates. Target-engaged CARs remain on proximal daughters, which inherit a surface proteome resembling activated-undivided CARTs, whereas the endogenous T cell receptor and CD8 enrich on distal daughters, whose surface proteome resembles resting CARTs, correlating with glycolytic and oxidative metabolism, respectively. Despite memory-precursor phenotype and in vivo longevity, distal daughters demonstrate transient potent cytolytic activity similar to proximal daughters, uncovering an effector-like state in distal daughters destined to become memory CARTs. Both partitioning of pre-existing transcripts and changes in RNA velocity contribute to asymmetry of fate-determining factors, resulting in diametrically opposed transcriptional trajectories. Independent of naive, memory or effector surface immunophenotype, proximal-daughter CARTs use core sets of transcription factors known to support proliferation and effector function. Conversely, transcription factors enriched in distal daughters restrain differentiation and promote longevity, evidenced by diminished long-term in vivo persistence and function of distal-daughter CARTs after IKZF1 disruption. These studies establish asymmetric cell division as a framework for understanding mechanisms of CART differentiation and improving therapeutic outcomes.
早期扩增和长期持续与嵌合抗原受体 T 细胞 (CART) 的疗效相关,但调节效应器与记忆性 CART 分化的机制,以及不对称细胞分裂是否会诱导人 CART 产生不同命运,这些仍不清楚。在这里,我们展示了靶标诱导的邻近标记可用于分离第一次有丝分裂的近端和远端 CD8 CART,这些 CART 不对称地分配其表面蛋白质组和转录组,导致不同的命运。靶标结合的 CAR 仍然存在于近端子细胞上,这些子细胞继承了类似于激活未分裂 CART 的表面蛋白质组,而内源性 T 细胞受体和 CD8 则富集在远端子细胞上,其表面蛋白质组类似于静止的 CART,分别与糖酵解和氧化代谢相关。尽管具有记忆前体表型和体内长期存活能力,但远端子细胞表现出类似于近端子细胞的短暂强大的细胞毒性活性,揭示了注定成为记忆 CART 的远端子细胞中的效应样状态。既有预先存在的转录本的分配,也有 RNA 速度的变化,这两者都有助于命运决定因素的不对称,从而导致完全相反的转录轨迹。不依赖于幼稚、记忆或效应表面免疫表型,近端子细胞使用已知支持增殖和效应功能的核心转录因子集。相反,在远端子细胞中富集的转录因子抑制分化并促进长寿,这表现在 IKZF1 破坏后,远端子细胞的长期体内持久性和功能降低。这些研究确立了不对称细胞分裂作为理解 CART 分化机制和改善治疗效果的框架。