VIB-KULeuven Centre for Cancer Biology, Department of Oncology, Leuven, Belgium.
Division of Molecular Pathology, Oncode Institute, The Netherlands Cancer Institute, Amsterdam, the Netherlands.
Nature. 2024 Sep;633(8028):198-206. doi: 10.1038/s41586-024-07882-3. Epub 2024 Sep 4.
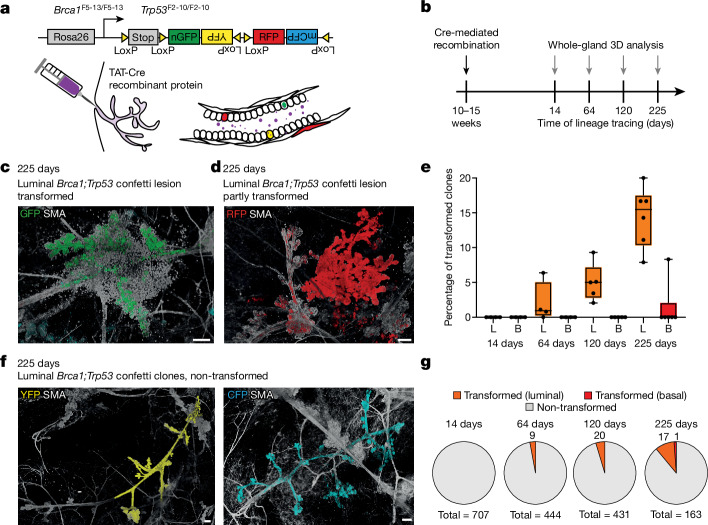
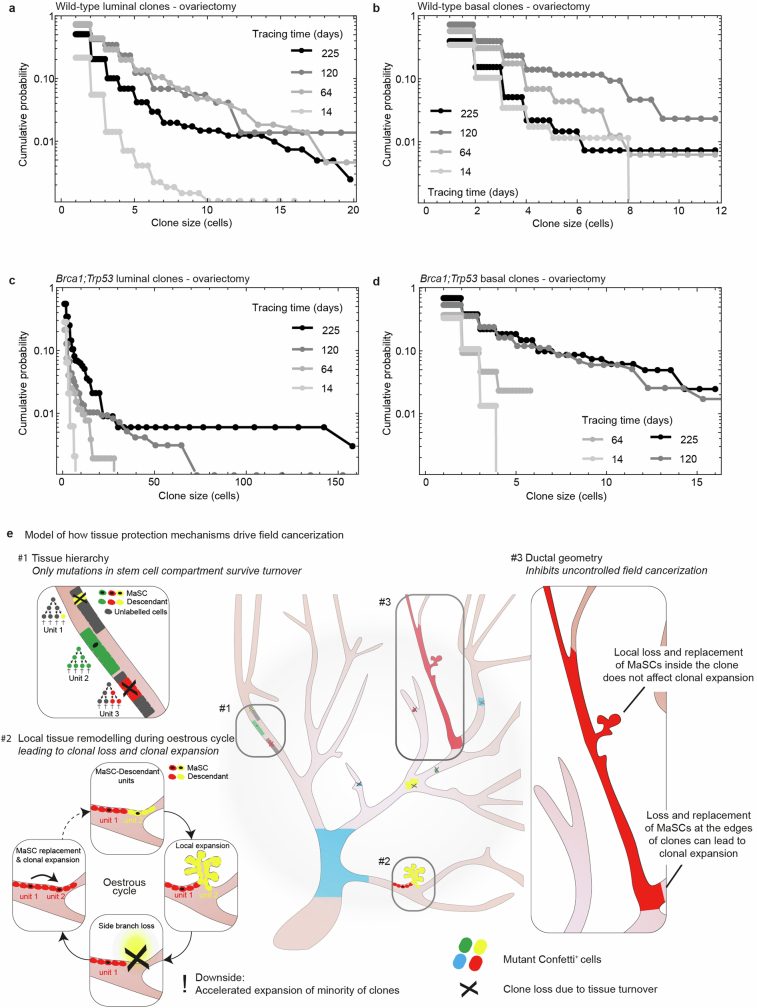
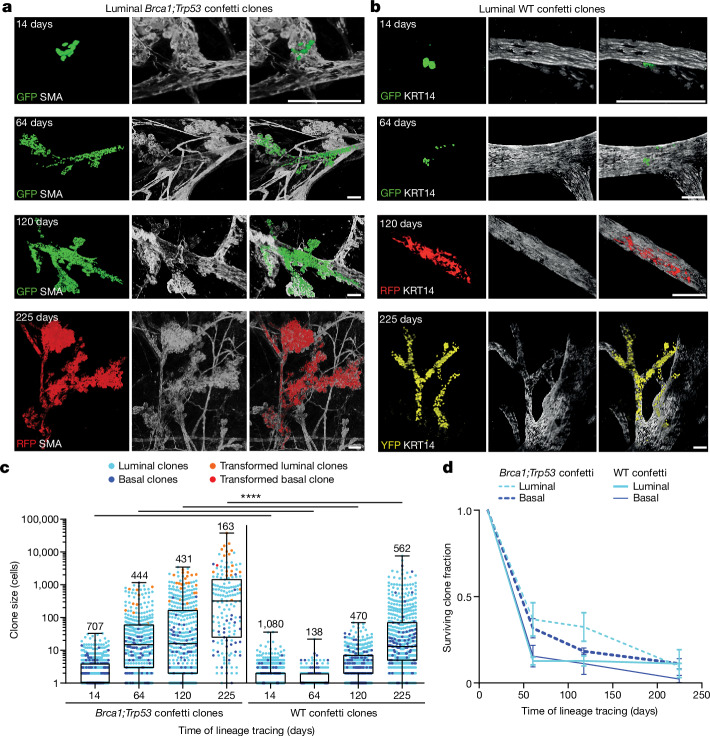
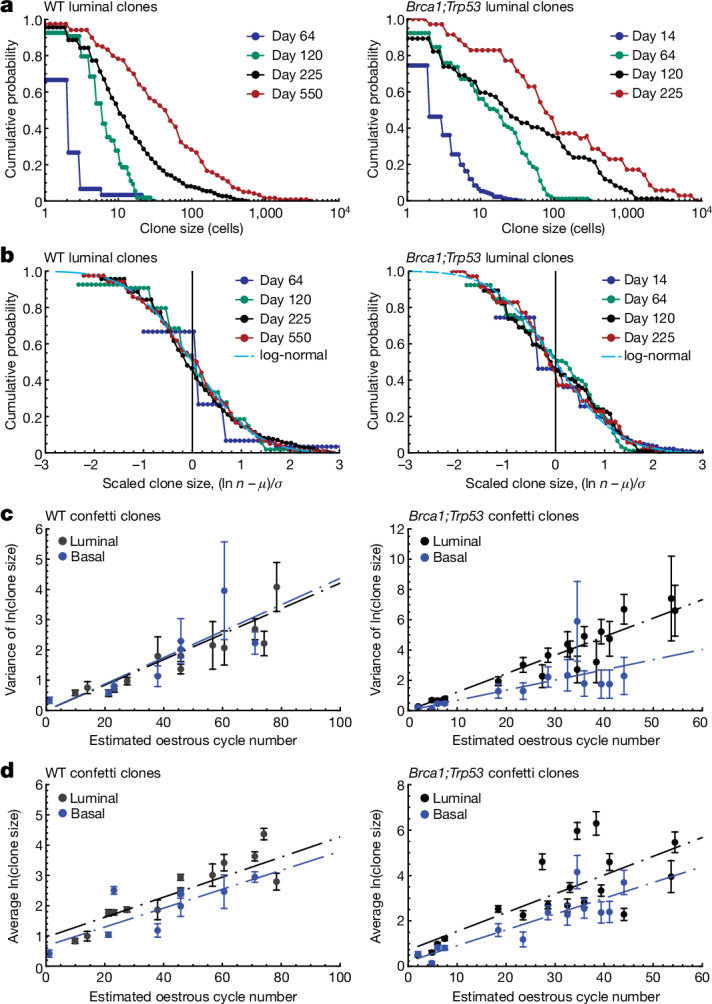
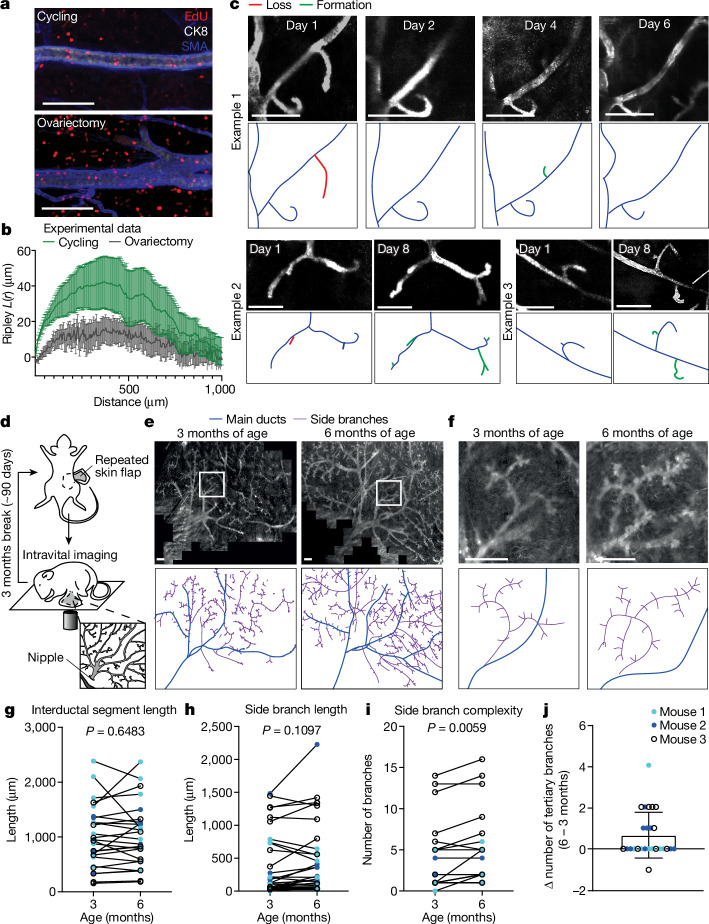
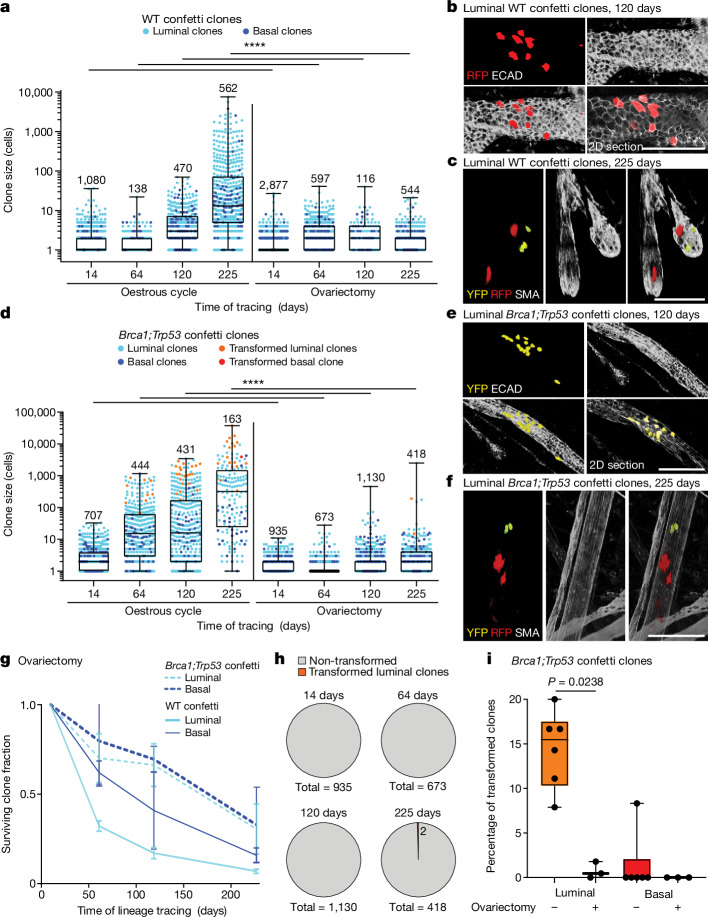
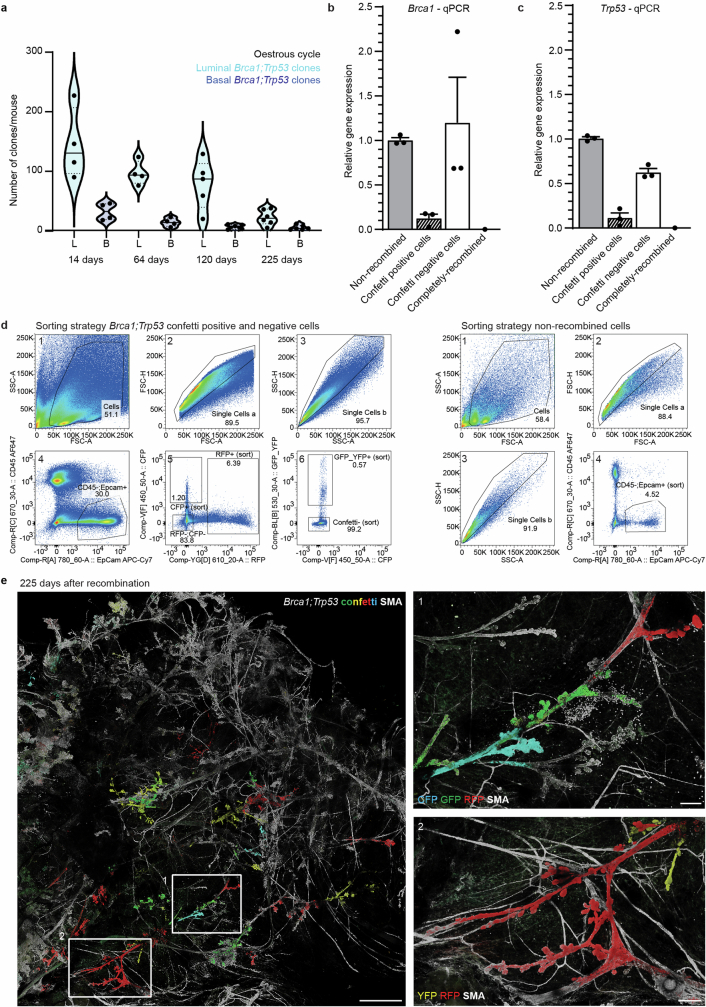
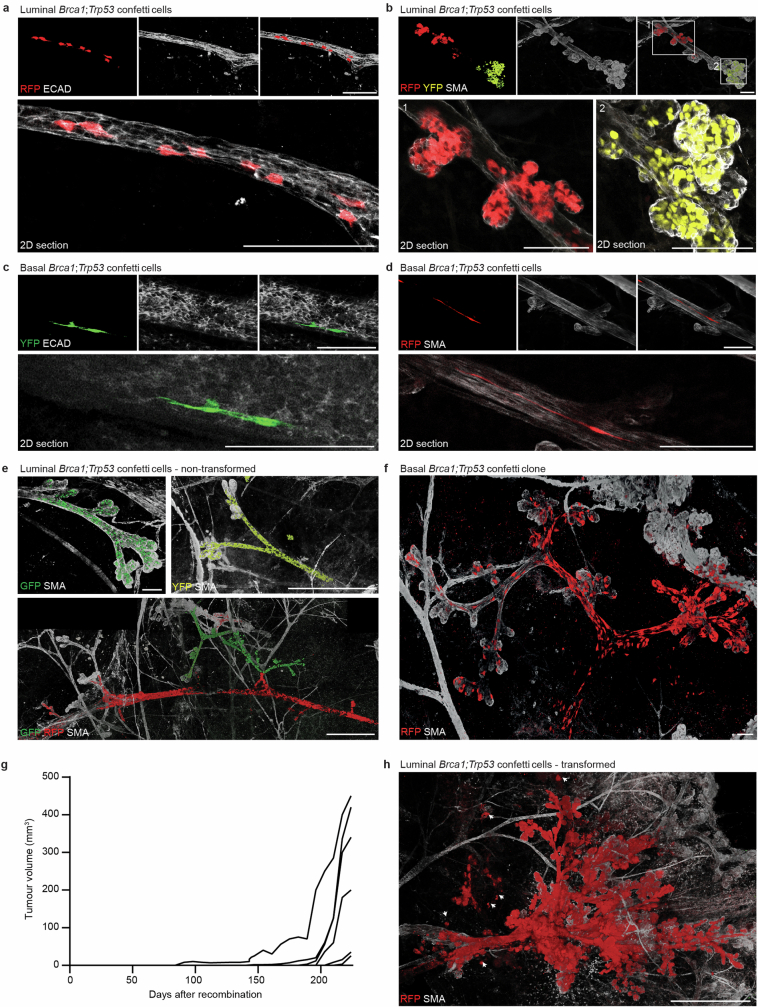
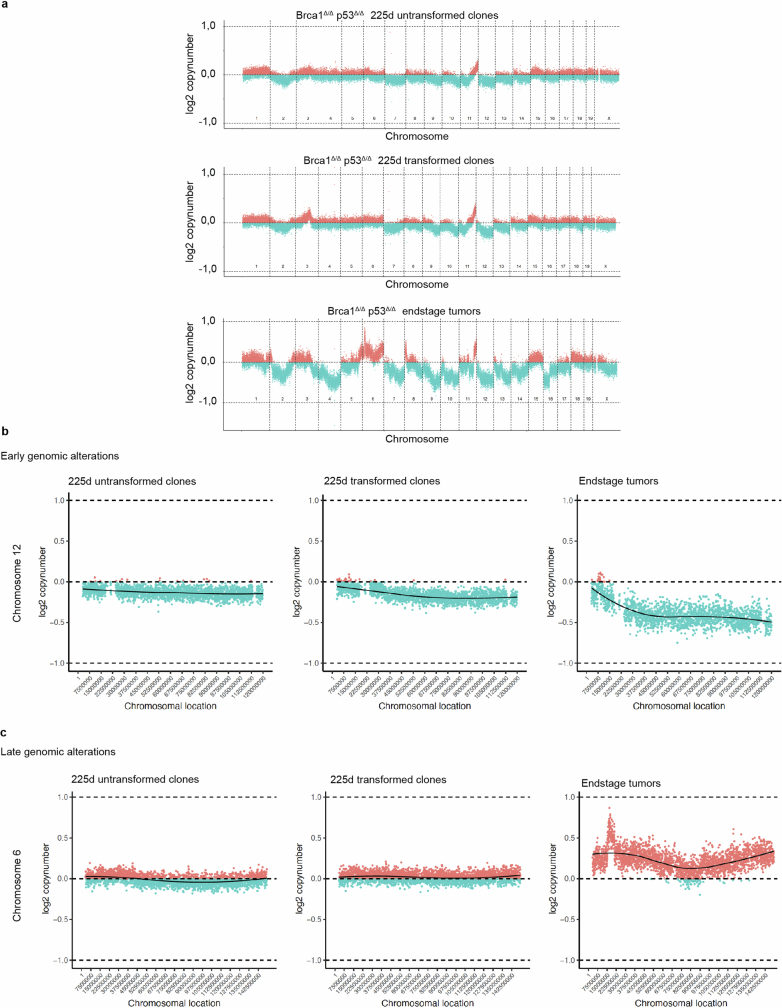
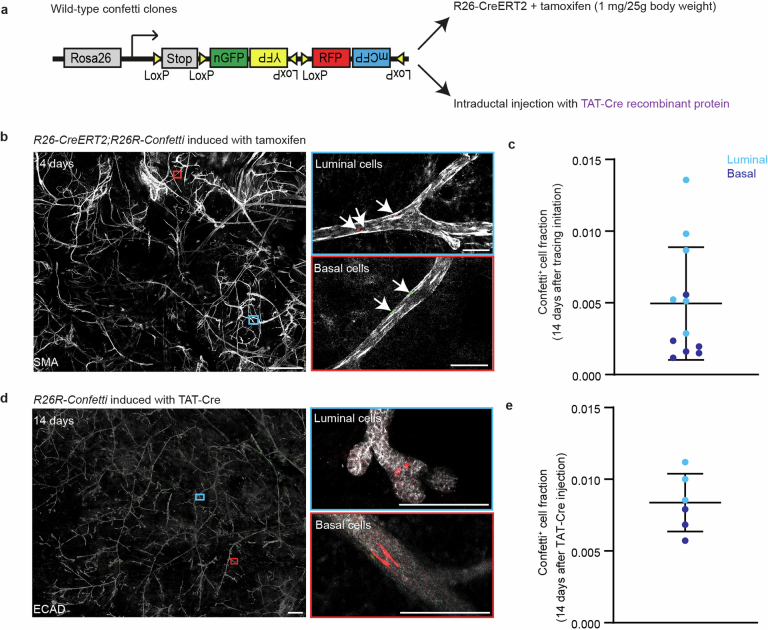
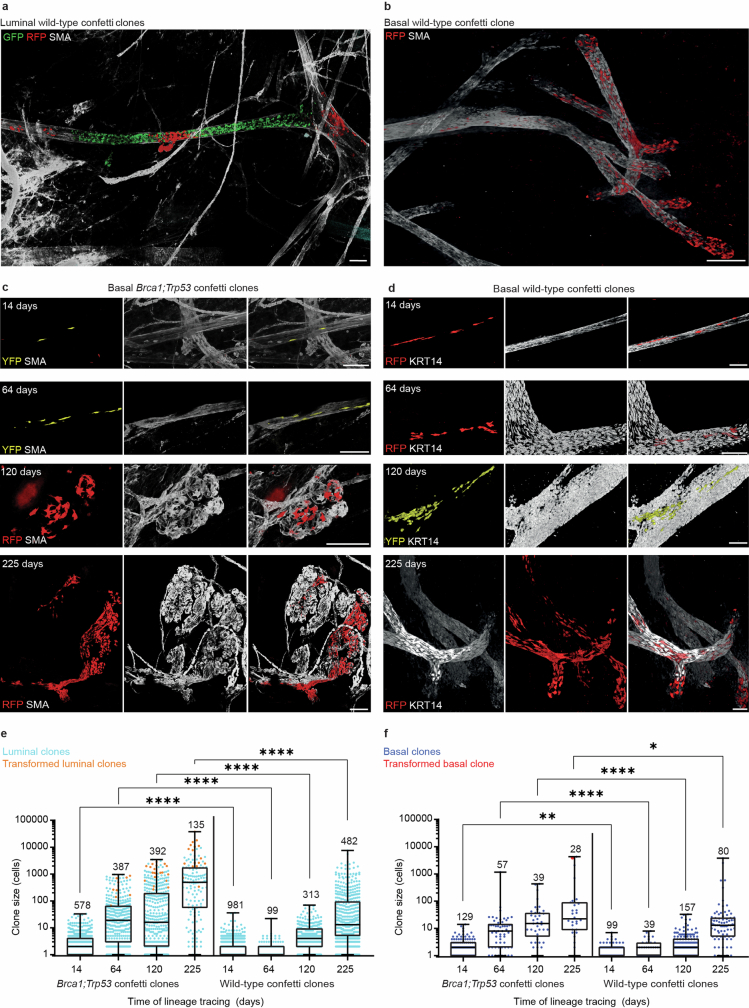
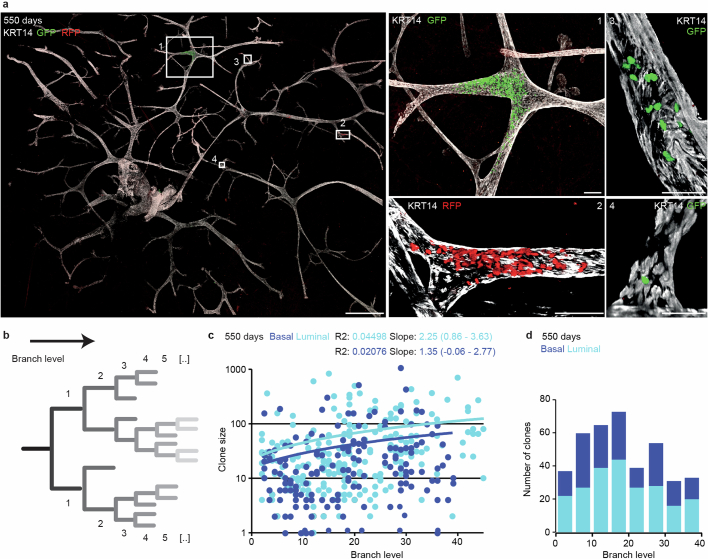
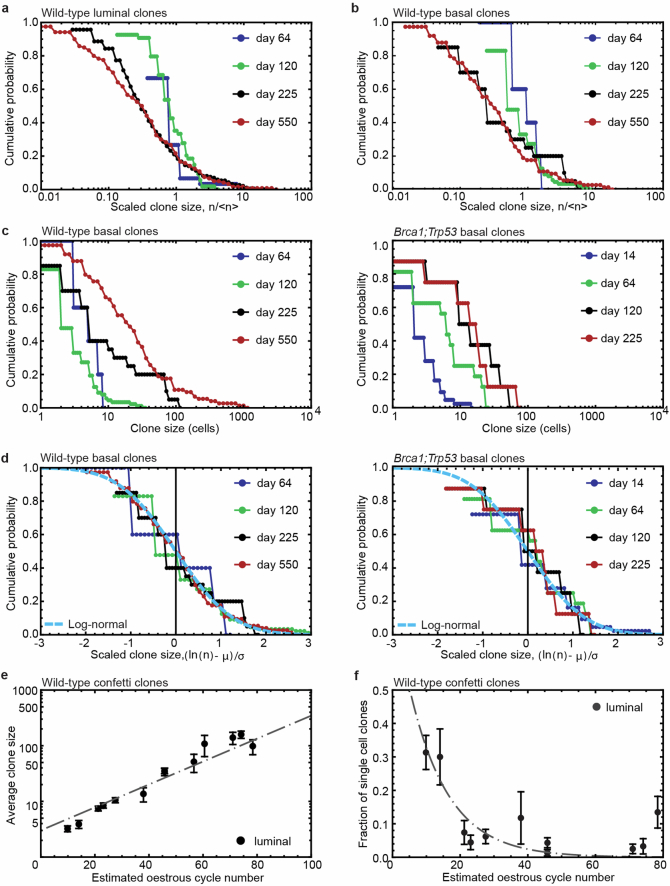
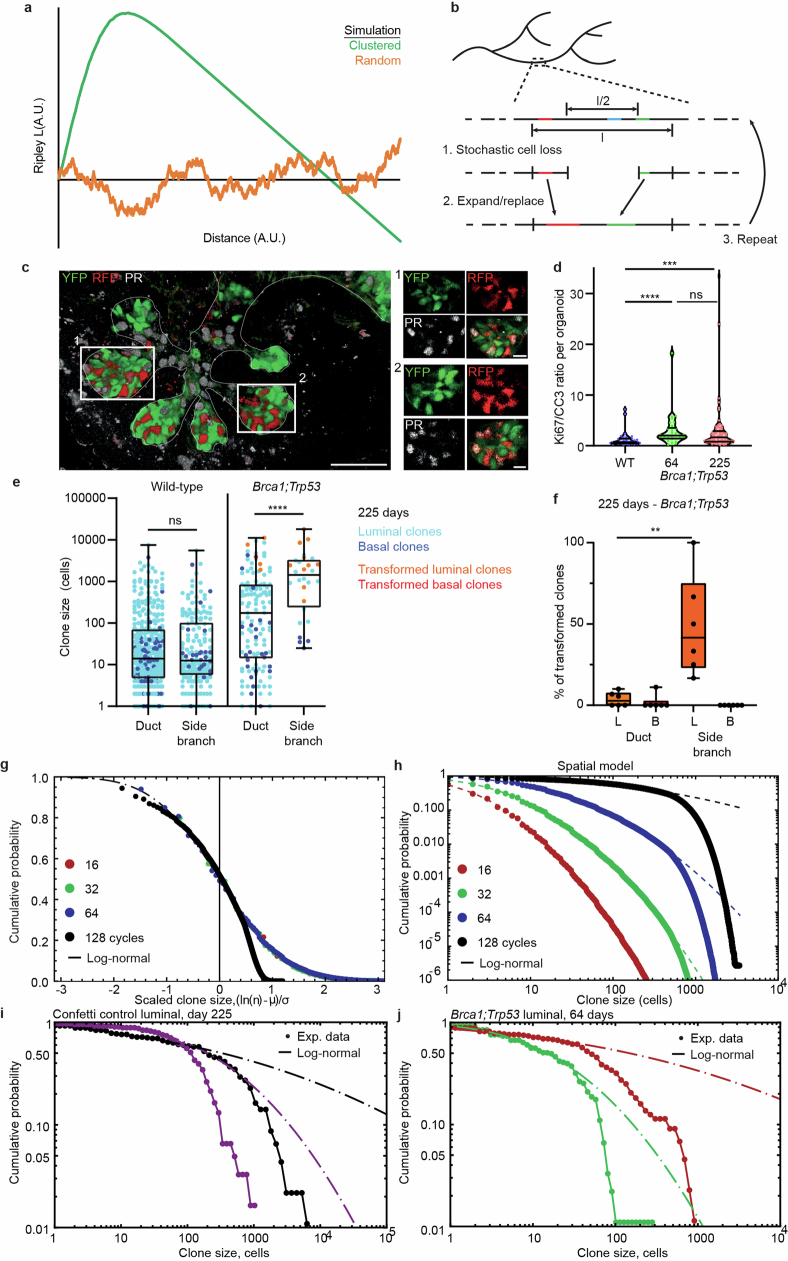
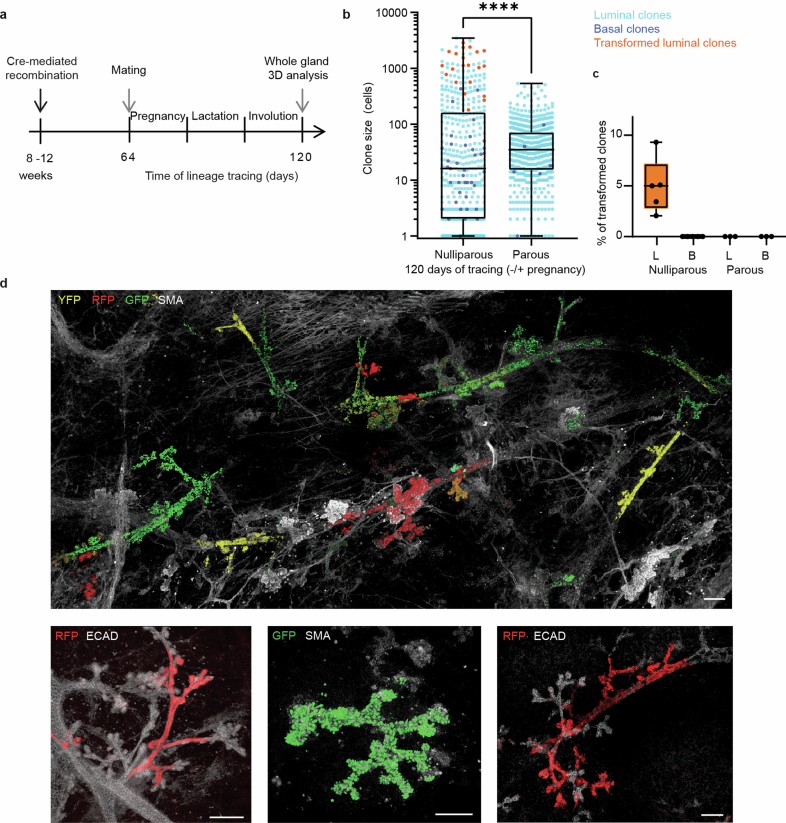
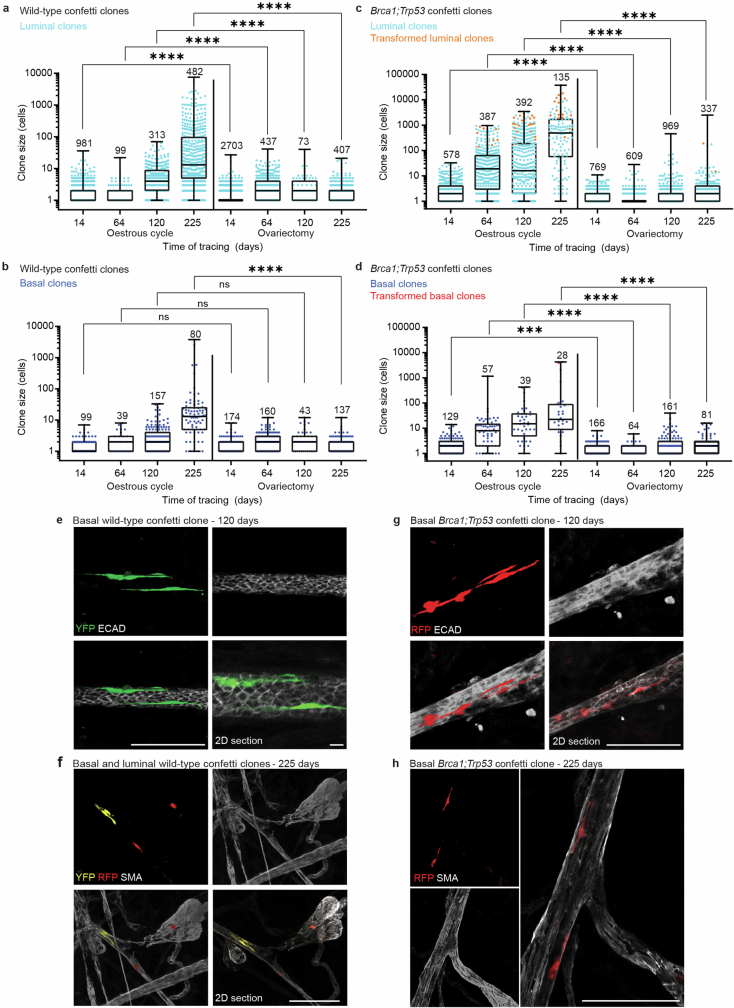
Oncogenic mutations are abundant in the tissues of healthy individuals, but rarely form tumours. Yet, the underlying protection mechanisms are largely unknown. To resolve these mechanisms in mouse mammary tissue, we use lineage tracing to map the fate of wild-type and Brca1;Trp53 cells, and find that both follow a similar pattern of loss and spread within ducts. Clonal analysis reveals that ducts consist of small repetitive units of self-renewing cells that give rise to short-lived descendants. This offers a first layer of protection as any descendants, including oncogenic mutant cells, are constantly lost, thereby limiting the spread of mutations to a single stem cell-descendant unit. Local tissue remodelling during consecutive oestrous cycles leads to the cooperative and stochastic loss and replacement of self-renewing cells. This process provides a second layer of protection, leading to the elimination of most mutant clones while enabling the minority that by chance survive to expand beyond the stem cell-descendant unit. This leads to fields of mutant cells spanning large parts of the epithelial network, predisposing it for transformation. Eventually, clone expansion becomes restrained by the geometry of the ducts, providing a third layer of protection. Together, these mechanisms act to eliminate most cells that acquire somatic mutations at the expense of driving the accelerated expansion of a minority of cells, which can colonize large areas, leading to field cancerization.
致癌突变在健康个体的组织中很常见,但很少形成肿瘤。然而,潜在的保护机制在很大程度上尚不清楚。为了在小鼠乳腺组织中解析这些机制,我们使用谱系追踪来追踪野生型和 Brca1;Trp53 细胞的命运,发现它们都遵循类似的导管内丢失和扩散模式。克隆分析表明,导管由自我更新细胞的小重复单元组成,这些细胞产生短暂寿命的后代。这提供了第一层保护,因为任何后代,包括致癌突变细胞,都会不断丢失,从而将突变的传播限制在单个干细胞-后代单元内。在连续发情周期中局部组织重塑导致自我更新细胞的协同和随机丢失和替换。这个过程提供了第二层保护,导致大多数突变克隆被消除,同时使少数有机会存活下来的克隆能够扩展到干细胞-后代单元之外。这导致跨越上皮网络大部分区域的突变细胞区域,使其容易发生转化。最终,由于导管的几何形状,克隆扩展受到限制,从而提供了第三层保护。这些机制共同作用,以牺牲驱动少数细胞加速扩张为代价,消除大多数获得体细胞突变的细胞,这些细胞可以殖民大面积,导致癌前病变。