MRC Laboratory of Molecular Biology, Cambridge, UK.
Department of Brain and Neurosciences, Tokyo Metropolitan Institute of Medical Science, Tokyo, Japan.
Nature. 2024 Oct;634(8034):662-668. doi: 10.1038/s41586-024-08024-5. Epub 2024 Sep 11.
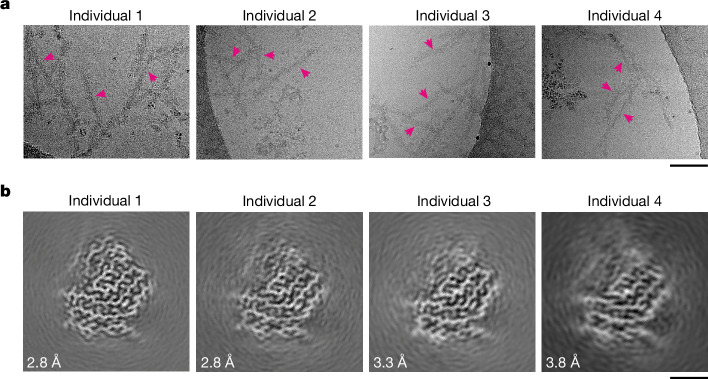
Neurodegenerative diseases are characterized by the abnormal filamentous assembly of specific proteins in the central nervous system. Human genetic studies have established a causal role for protein assembly in neurodegeneration. However, the underlying molecular mechanisms remain largely unknown, which is limiting progress in developing clinical tools for these diseases. Recent advances in cryo-electron microscopy have enabled the structures of the protein filaments to be determined from the brains of patients. All neurodegenerative diseases studied to date have been characterized by the self-assembly of proteins in homomeric amyloid filaments, including that of TAR DNA-binding protein 43 (TDP-43) in amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration with TDP-43 inclusions (FTLD-TDP) types A and B. Here we used cryo-electron microscopy to determine filament structures from the brains of individuals with FTLD-TDP type C, one of the most common forms of sporadic FTLD-TDP. Unexpectedly, the structures revealed that a second protein, annexin A11 (ANXA11), co-assembles with TDP-43 in heteromeric amyloid filaments. The ordered filament fold is formed by TDP-43 residues G282/G284-N345 and ANXA11 residues L39-Y74 from their respective low-complexity domains. Regions of TDP-43 and ANXA11 that were previously implicated in protein-protein interactions form an extensive hydrophobic interface at the centre of the filament fold. Immunoblots of the filaments revealed that the majority of ANXA11 exists as an approximately 22 kDa N-terminal fragment lacking the annexin core domain. Immunohistochemistry of brain sections showed the colocalization of ANXA11 and TDP-43 in inclusions, redefining the histopathology of FTLD-TDP type C. This work establishes a central role for ANXA11 in FTLD-TDP type C. The unprecedented formation of heteromeric amyloid filaments in the human brain revises our understanding of amyloid assembly and may be of significance for the pathogenesis of neurodegenerative diseases.
神经退行性疾病的特征是中枢神经系统中特定蛋白质的异常丝状组装。人类遗传研究已经确定了蛋白质组装在神经退行性变中的因果作用。然而,其潜在的分子机制在很大程度上仍然未知,这限制了开发这些疾病的临床工具的进展。最近的低温电子显微镜技术的进步使得能够从患者的大脑中确定蛋白质丝的结构。迄今为止,所有研究的神经退行性疾病都表现为同种蛋白在同源淀粉样纤维中的自组装,包括肌萎缩侧索硬化症(ALS)和额颞叶痴呆伴 TDP-43 包涵体(FTLD-TDP)的 TAR DNA 结合蛋白 43(TDP-43)。在这里,我们使用低温电子显微镜来确定来自 FTLD-TDP 类型 C 个体的纤维结构,这是最常见的散发性 FTLD-TDP 形式之一。出乎意料的是,结构表明第二种蛋白质,膜联蛋白 A11(ANXA11)与 TDP-43 异源淀粉样纤维中组装。由 TDP-43 的残基 G282/G284-N345 和 ANXA11 的残基 L39-Y74 从其各自的低复杂度结构域形成有序的纤维折叠。先前涉及蛋白质-蛋白质相互作用的 TDP-43 和 ANXA11 的区域在纤维折叠的中心形成一个广泛的疏水区。纤维的免疫印迹显示,大多数 ANXA11 以缺乏膜联蛋白核心结构域的约 22 kDa N 端片段形式存在。脑切片的免疫组织化学显示 ANXA11 和 TDP-43 在包涵体中的共定位,重新定义了 FTLD-TDP 类型 C 的组织病理学。这项工作确立了 ANXA11 在 FTLD-TDP 类型 C 中的核心作用。在人类大脑中前所未有的异源淀粉样纤维的形成改变了我们对淀粉样纤维组装的理解,可能对神经退行性疾病的发病机制具有重要意义。