Department of Molecular, Cellular, and Developmental Biology, University of Michigan, Ann Arbor, Michigan, United States of America.
PLoS Biol. 2024 Sep 24;22(9):e3002368. doi: 10.1371/journal.pbio.3002368. eCollection 2024 Sep.
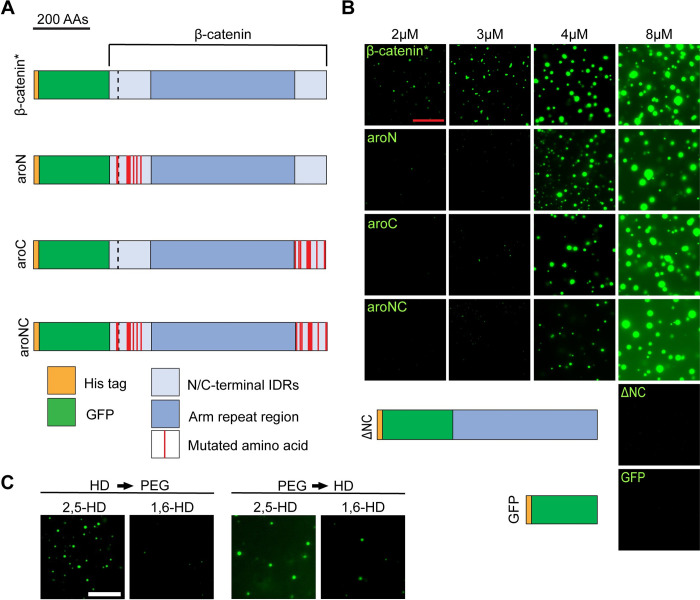
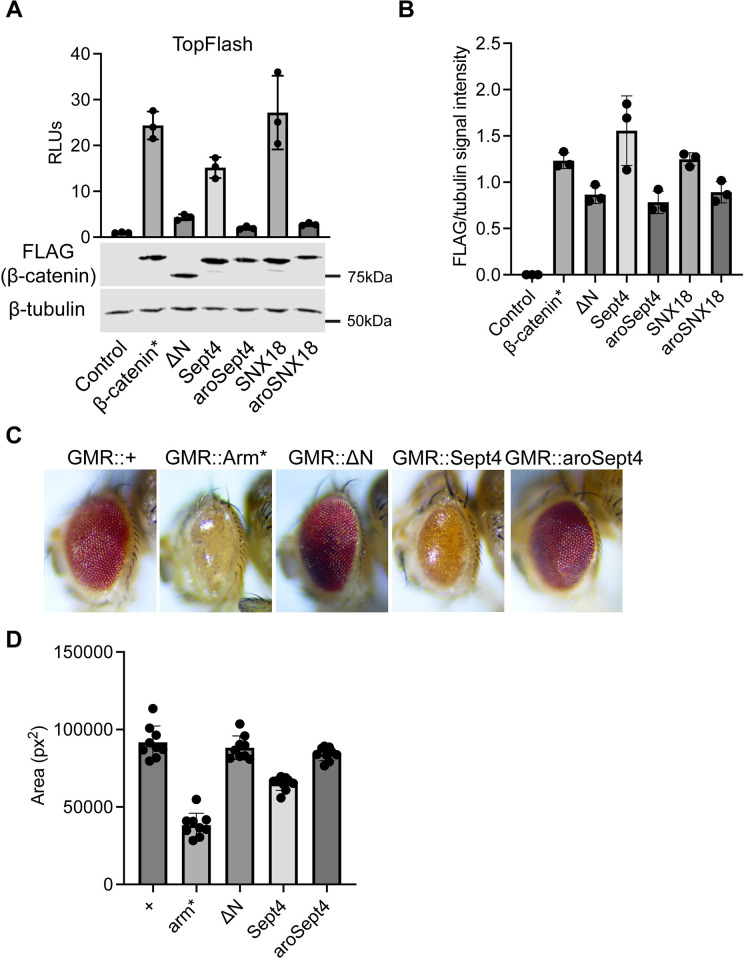
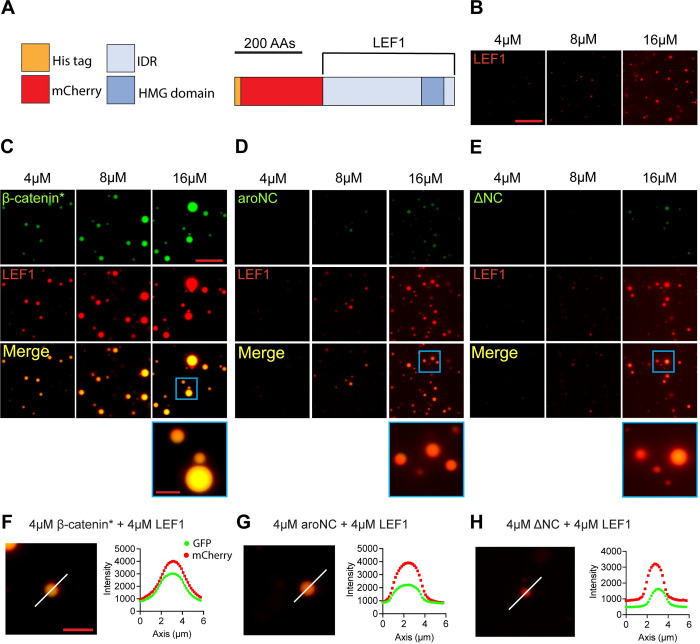
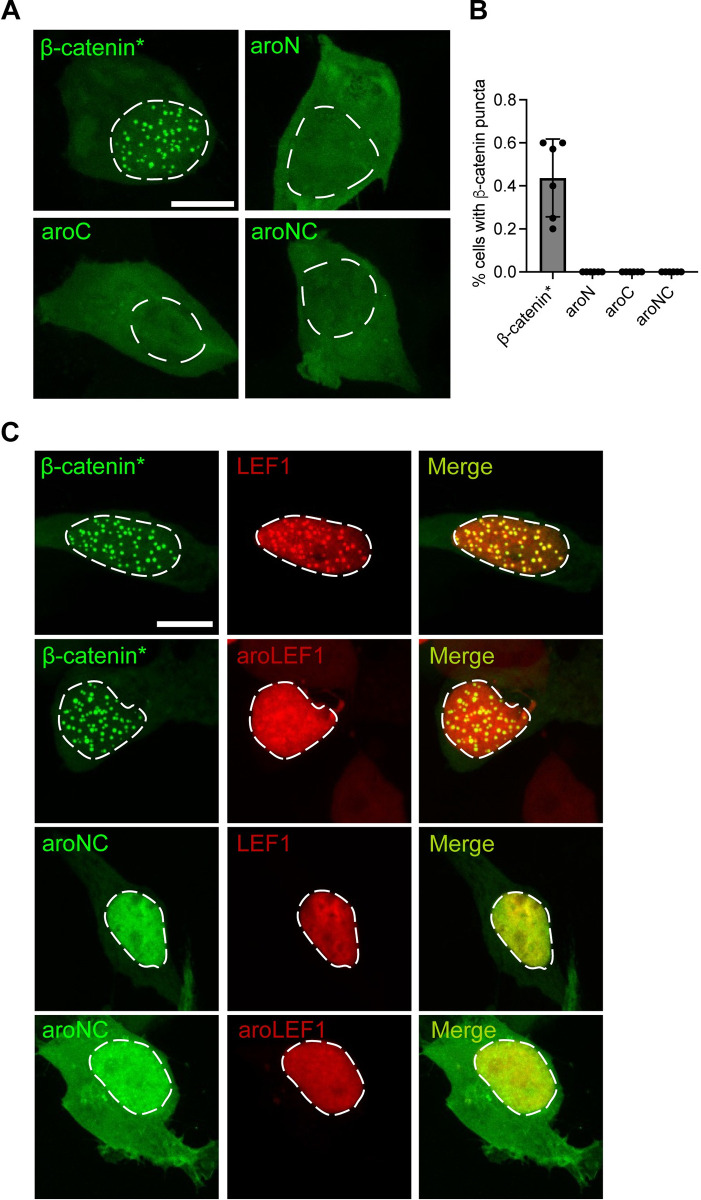
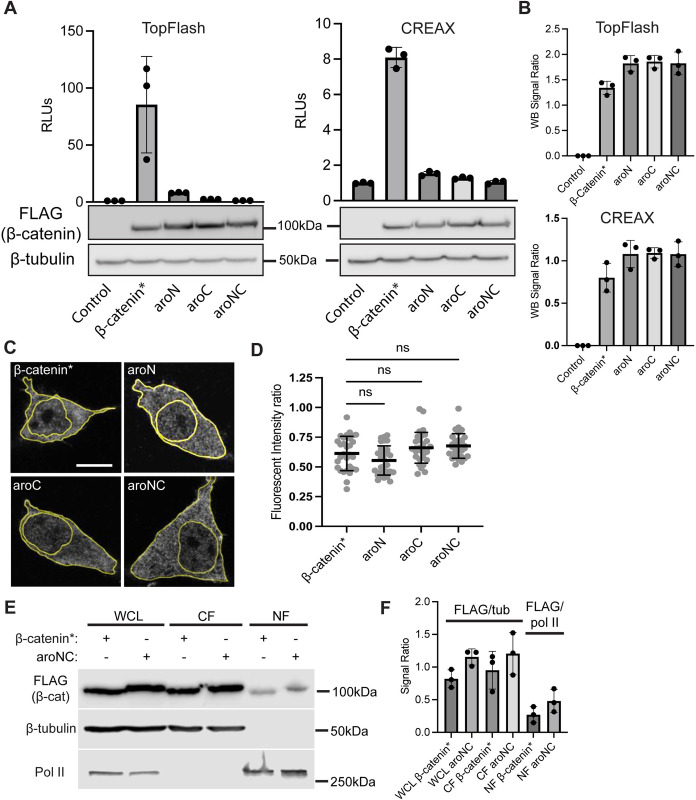
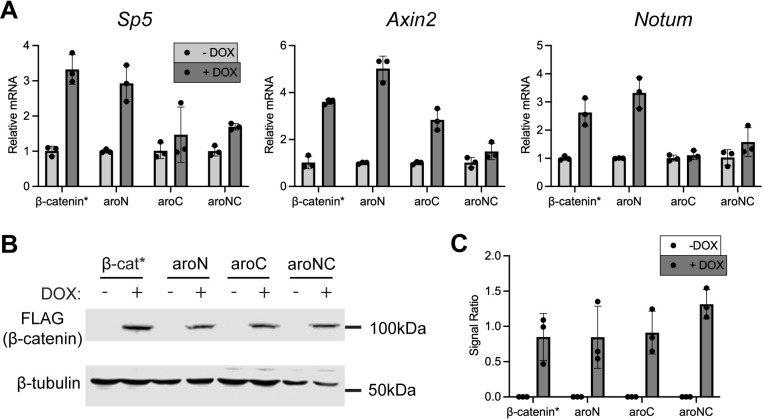
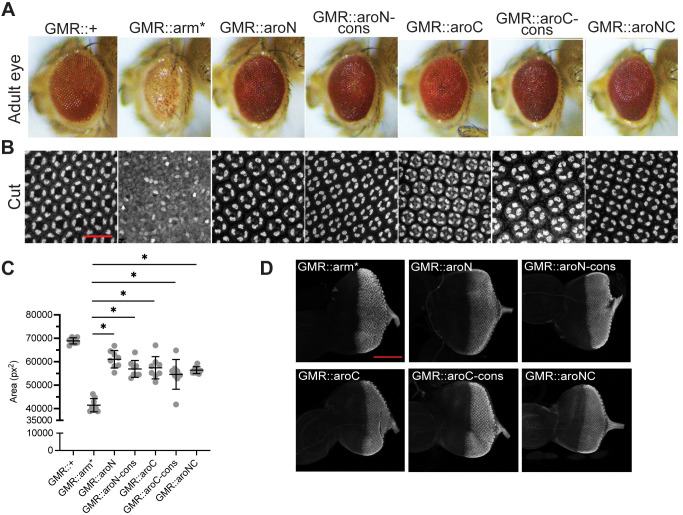
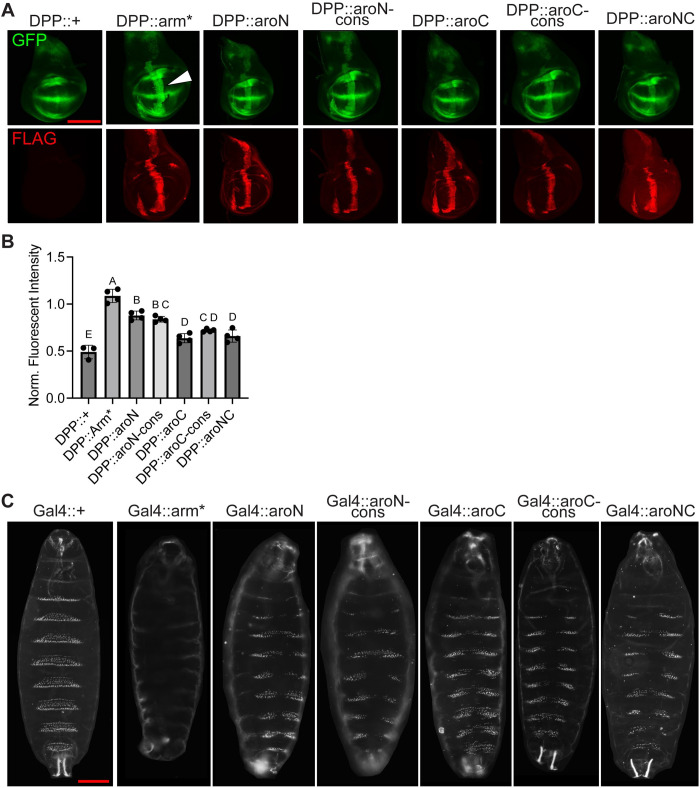
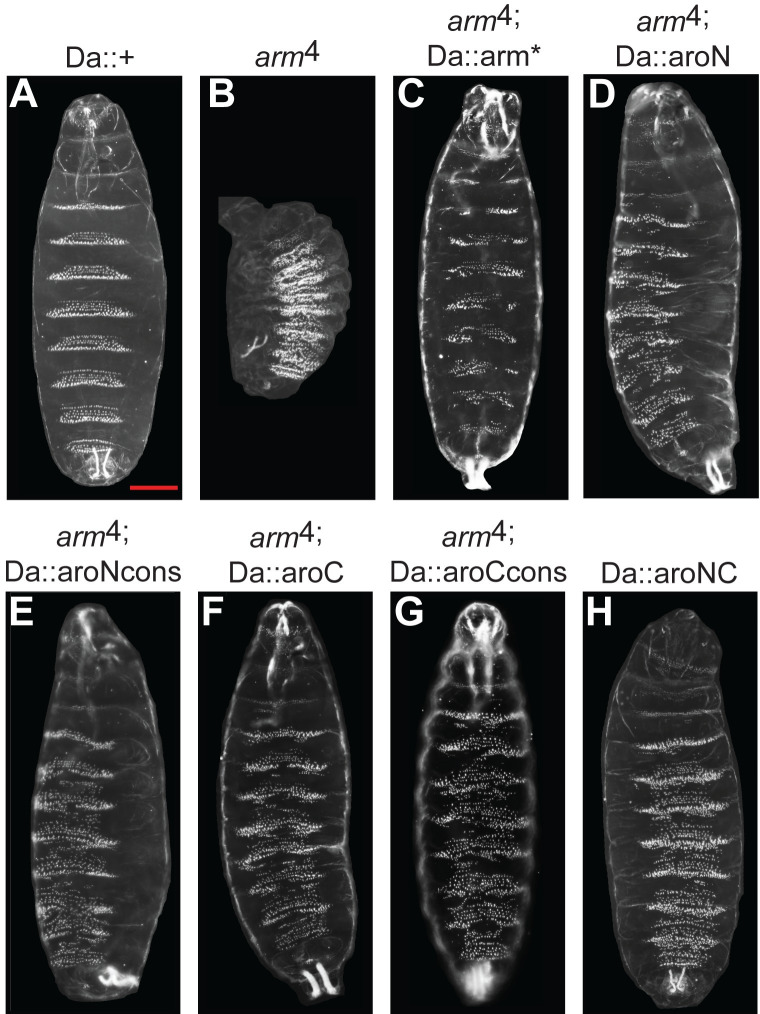
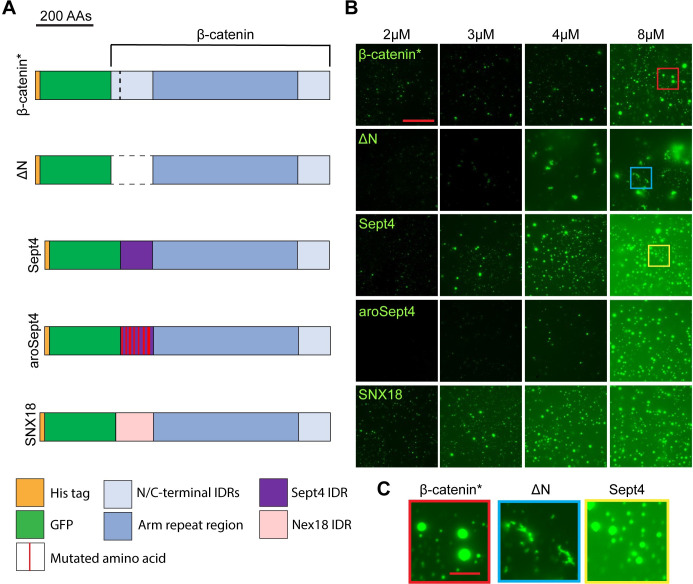
The Wnt/β-catenin signaling pathway plays numerous essential roles in animal development and tissue/stem cell maintenance. The activation of genes regulated by Wnt/β-catenin signaling requires the nuclear accumulation of β-catenin, a transcriptional co-activator. β-catenin is recruited to many Wnt-regulated enhancers through direct binding to T-cell factor/lymphoid enhancer factor (TCF/LEF) family transcription factors. β-catenin has previously been reported to form phase-separated biomolecular condensates (BMCs), which was implicated as a component of β-catenin's mechanism of action. This function required aromatic amino acid residues in the intrinsically disordered regions (IDRs) at the N- and C-termini of the protein. In this report, we further explore a role for β-catenin BMCs in Wnt target gene regulation. We find that β-catenin BMCs are miscible with LEF1 BMCs in vitro and in cultured cells. We characterized a panel of β-catenin mutants with different combinations of aromatic residue mutations in human cell culture and Drosophila melanogaster. Our data support a model in which aromatic residues across both IDRs contribute to BMC formation and signaling activity. Although different Wnt targets have different sensitivities to loss of β-catenin's aromatic residues, the activation of every target examined was compromised by aromatic substitution. These mutants are not defective in nuclear import or co-immunoprecipitation with several β-catenin binding partners. In addition, residues in the N-terminal IDR with no previously known role in signaling are clearly required for the activation of various Wnt readouts. Consistent with this, deletion of the N-terminal IDR results in a loss of signaling activity, which can be rescued by the addition of heterologous IDRs enriched in aromatic residues. Overall, our work supports a model in which the ability of β-catenin to form biomolecular condensates in the nucleus is tightly linked to its function as a transcriptional co-regulator.
Wnt/β-catenin 信号通路在动物发育和组织/干细胞维持中发挥着许多重要作用。Wnt/β-catenin 信号的激活需要β-连环蛋白(β-catenin)的核积累,β-catenin 是一种转录共激活因子。β-catenin 通过直接结合 T 细胞因子/淋巴增强因子(TCF/LEF)家族转录因子,被招募到许多受 Wnt 调节的增强子上。β-catenin 先前被报道形成相分离的生物分子凝聚物(BMCs),这被认为是 β-catenin 作用机制的一个组成部分。该功能需要蛋白质 N 端和 C 端无规卷曲区域(IDR)中的芳香族氨基酸残基。在本报告中,我们进一步探讨了β-catenin BMCs 在 Wnt 靶基因调控中的作用。我们发现β-catenin BMCs 在体外和培养细胞中与 LEF1 BMCs 可混溶。我们在人细胞培养和黑腹果蝇中表征了一组具有不同芳香族残基突变组合的β-catenin 突变体。我们的数据支持这样一种模型,即 IDR 两端的芳香族残基有助于 BMC 形成和信号转导活性。尽管不同的 Wnt 靶标对β-catenin 的芳香族残基缺失的敏感性不同,但所研究的每个靶标的激活都因芳香族取代而受损。这些突变体在核输入或与几个β-catenin 结合伙伴的共免疫沉淀中没有缺陷。此外,在信号传导中没有先前已知作用的 N 端 IDR 中的残基对于各种 Wnt 读出物的激活显然是必需的。与这一点一致的是,N 端 IDR 的缺失导致信号转导活性丧失,而添加富含芳香族残基的异源 IDR 可以挽救该活性。总的来说,我们的工作支持这样一种模型,即β-catenin 在核内形成生物分子凝聚物的能力与其作为转录共调节因子的功能紧密相关。