Department of Cell Biology, Faculty of Medicine, Fukuoka University, 7-45-1 Nanakuma, Jonan-ku, Fukuoka 814-0180, Japan.
Department of Anesthesiology, Faculty of Medicine, Fukuoka University, 7-45-1 Nanakuma, Jonan-ku, Fukuoka 814-0180, Japan.
Nucleic Acids Res. 2024 Nov 27;52(21):12961-12975. doi: 10.1093/nar/gkae852.
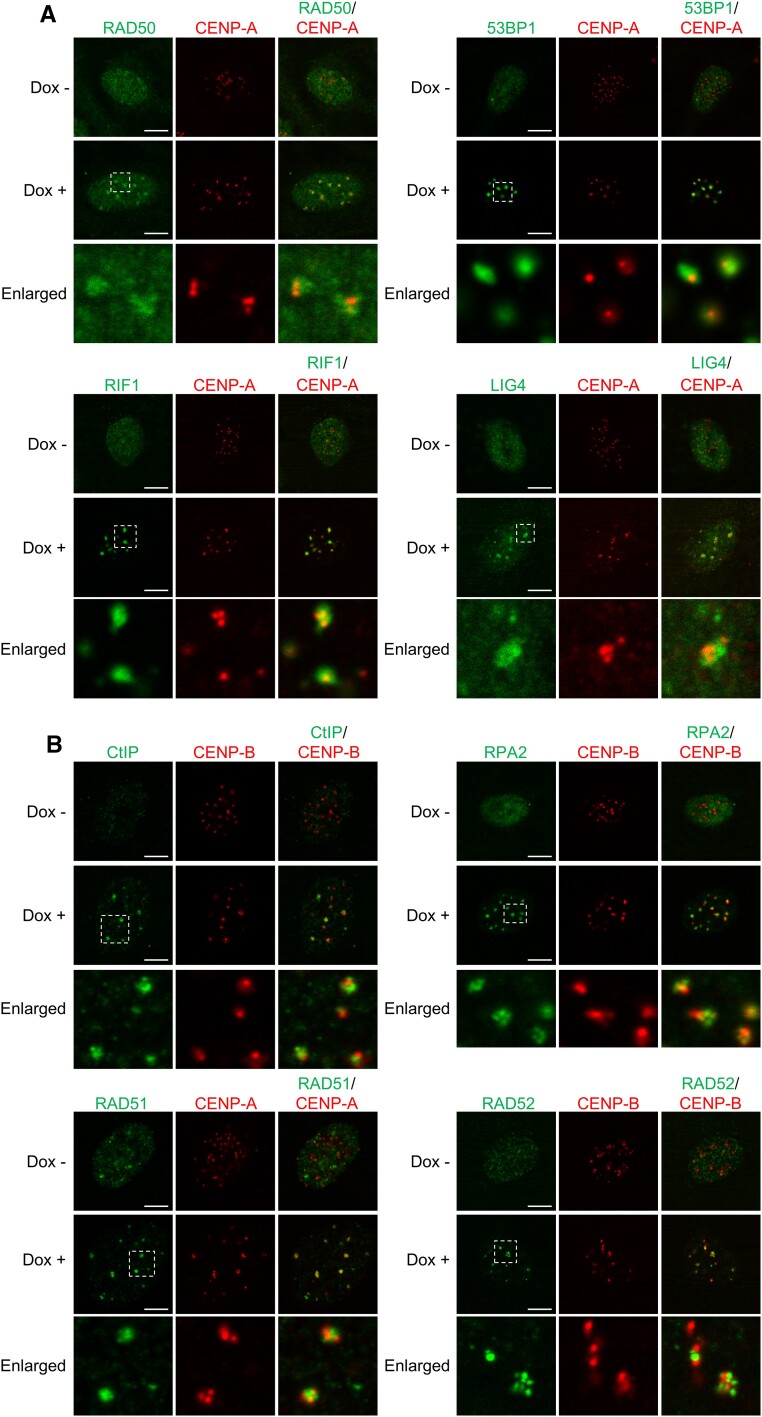
Centromeres, where the kinetochore complex binds, are susceptible to damages including DNA double-stranded breaks (DSBs). Here, we report the functional significance and the temporally and spatially distinct regulation of centromeric DSB repair via the three pathways of non-homologous end joining (NHEJ), homologous recombination (HR) and single-strand annealing (SSA). The SSA factor RAD52 is most frequently recruited to centromeric DSB sites compared with the HR factor RAD51 and the NHEJ factor DNA ligase IV (LIG4), indicating that SSA plays predominant roles in centromeric DSB repair. Upon centromeric DSB induction, LIG4 is recruited to both active centromeres, where kinetochore complex binds, and inactive centromeres. In contrast, RAD51 and RAD52 are recruited only to inactive centromeres. These results indicate that DSBs at active centromeres are repaired through NHEJ, whereas the three pathways of NHEJ, HR and SSA are involved in DSB repair at inactive centromeres. Furthermore, siRNA-mediated depletion of either LIG4 or RAD51 promotes cell death after centromeric DSB induction, whereas RAD52 depletion inhibits it, suggesting that HR and NHEJ are required for appropriate centromeric DSB repair, whereas SSA-mediated centromeric DSB repair leads to subsequent cell death. Thus, SSA-mediated DSB repair at inactive centromeres may cause centromere dysfunction through error-prone repair.
着丝粒是动粒复合物结合的部位,容易受到包括 DNA 双链断裂 (DSB) 在内的损伤。在这里,我们报告了通过非同源末端连接 (NHEJ)、同源重组 (HR) 和单链退火 (SSA) 这三种途径修复着丝粒 DSB 的功能意义以及时空上的不同调控。与 HR 因子 RAD51 和 NHEJ 因子 DNA 连接酶 IV (LIG4) 相比,SSA 因子 RAD52 最常被招募到着丝粒 DSB 位点,这表明 SSA 在着丝粒 DSB 修复中发挥主要作用。在着丝粒 DSB 诱导后,LIG4 被招募到活跃的着丝粒(动粒复合物结合的部位)和非活跃的着丝粒。相比之下,RAD51 和 RAD52 仅被招募到非活跃的着丝粒。这些结果表明,活跃着丝粒上的 DSB 通过 NHEJ 修复,而 NHEJ、HR 和 SSA 这三种途径都参与了非活跃着丝粒上的 DSB 修复。此外,siRNA 介导的 LIG4 或 RAD51 耗竭会促进着丝粒 DSB 诱导后的细胞死亡,而 RAD52 耗竭则抑制细胞死亡,这表明 HR 和 NHEJ 是适当的着丝粒 DSB 修复所必需的,而 SSA 介导的着丝粒 DSB 修复会导致随后的细胞死亡。因此,非活跃着丝粒上的 SSA 介导的 DSB 修复可能会通过易错修复导致着丝粒功能障碍。