Mitra Sreyoshi, Gómez-Raja Jonathan, Larriba Germán, Dubey Dharani Dhar, Sanyal Kaustuv
Molecular Mycology Laboratory, Molecular Biology and Genetics Unit, Jawaharlal Nehru Centre for Advanced Scientific Research, Jakkur, Bangalore, India.
Departamento Ciencias Biomédicas Área de Microbiología, Universidad de Extremadura, Badajoz, Spain.
PLoS Genet. 2014 Apr 24;10(4):e1004344. doi: 10.1371/journal.pgen.1004344. eCollection 2014 Apr.
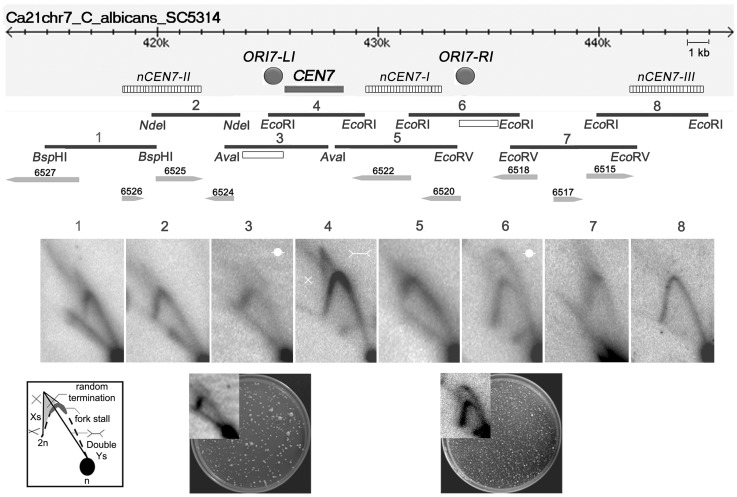
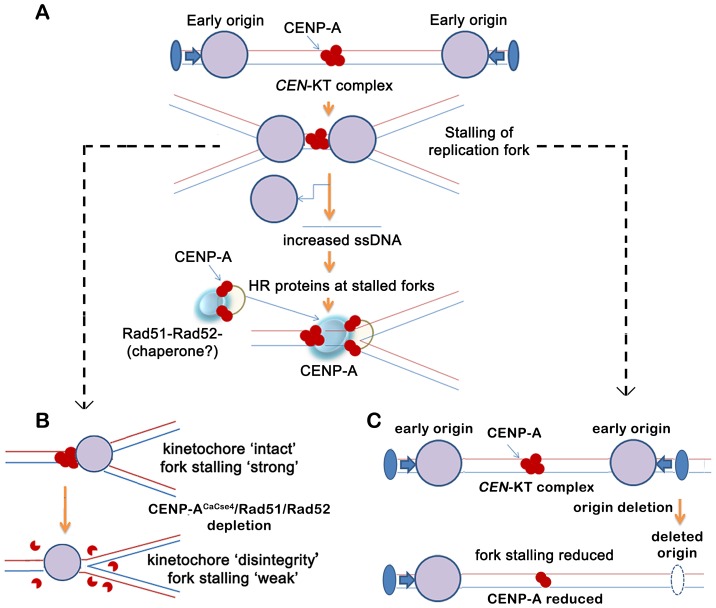
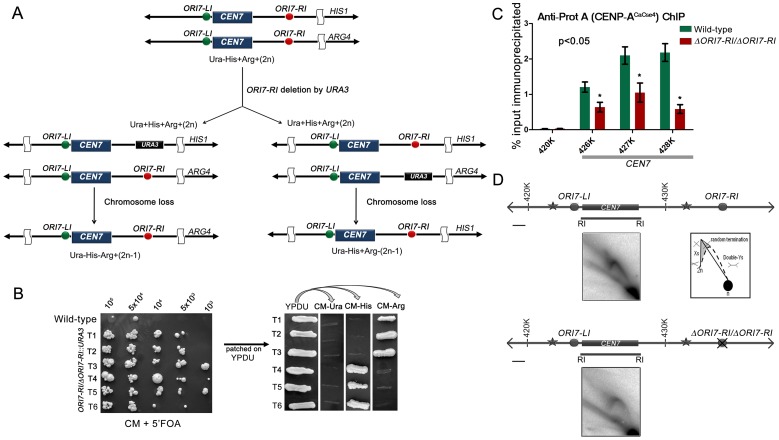
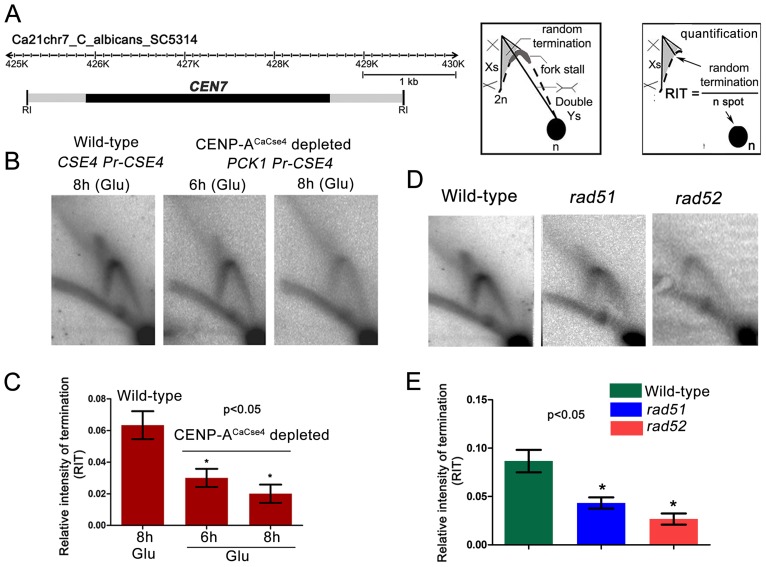
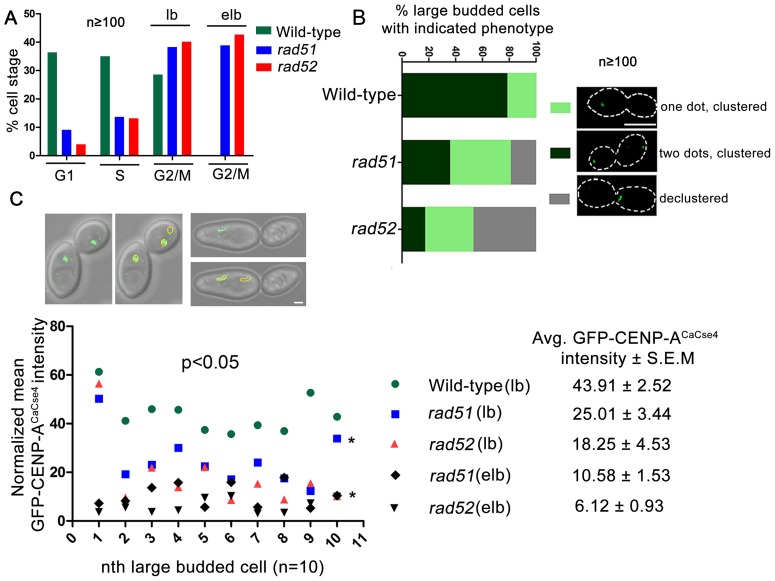
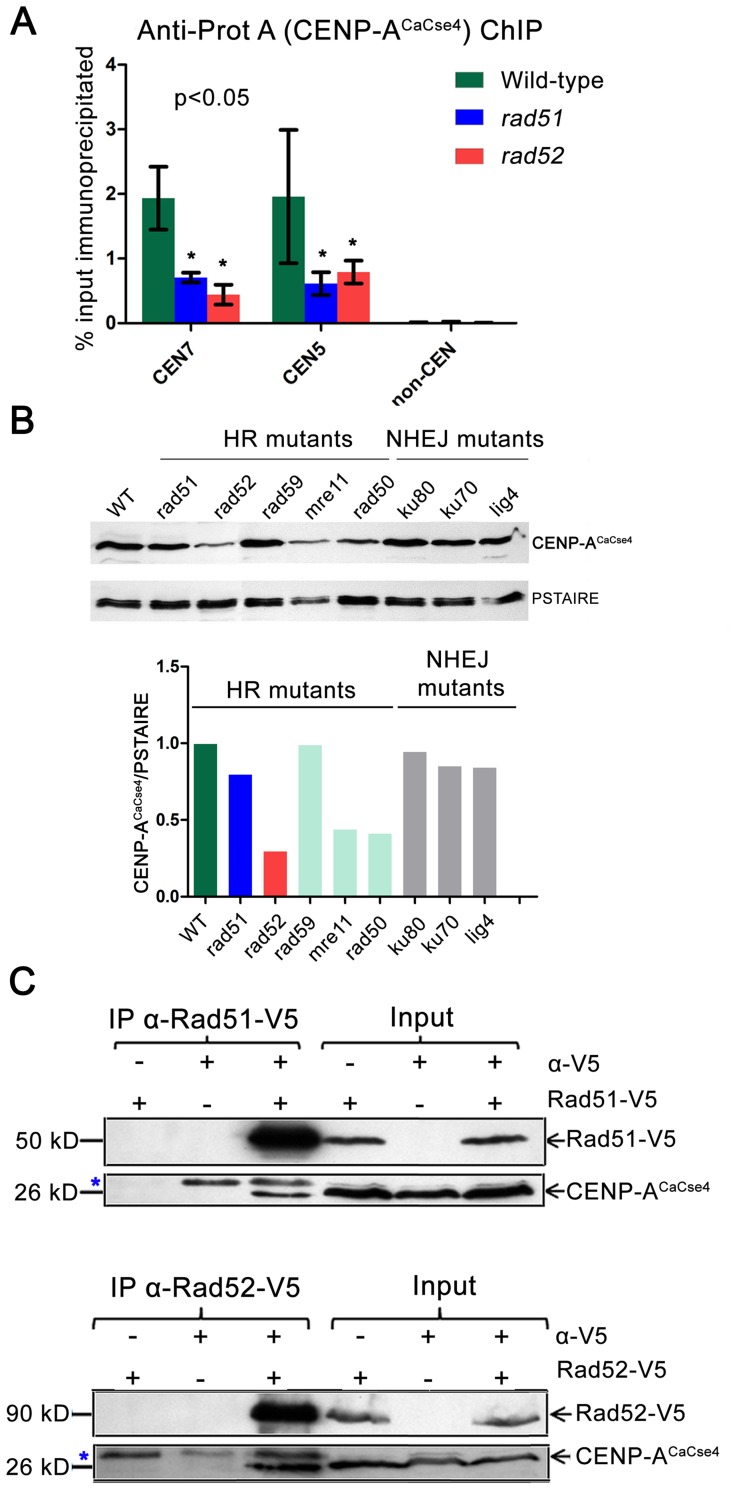
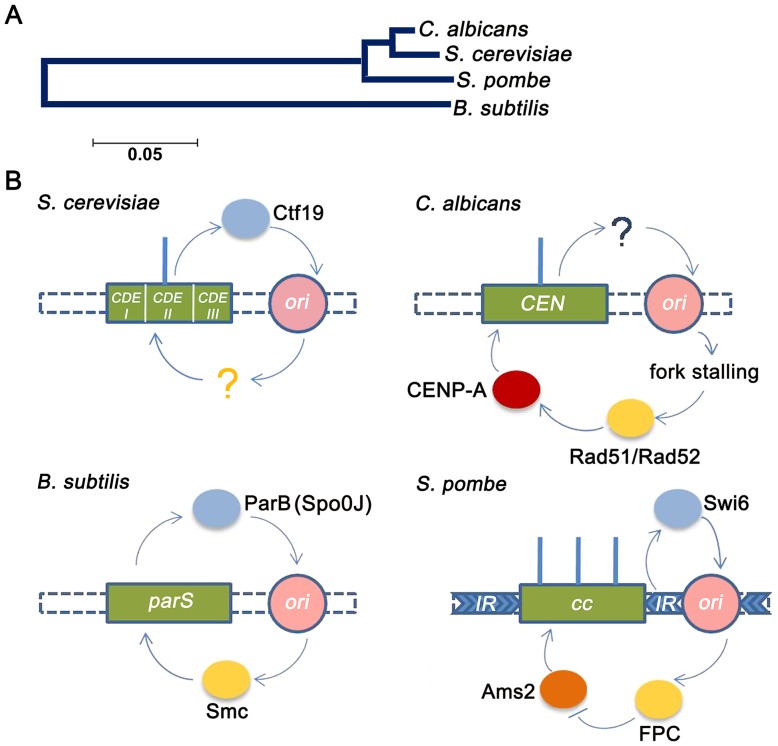
Specification of the centromere location in most eukaryotes is not solely dependent on the DNA sequence. However, the non-genetic determinants of centromere identity are not clearly defined. While multiple mechanisms, individually or in concert, may specify centromeres epigenetically, most studies in this area are focused on a universal factor, a centromere-specific histone H3 variant CENP-A, often considered as the epigenetic determinant of centromere identity. In spite of variable timing of its loading at centromeres across species, a replication coupled early S phase deposition of CENP-A is found in most yeast centromeres. Centromeres are the earliest replicating chromosomal regions in a pathogenic budding yeast Candida albicans. Using a 2-dimensional agarose gel electrophoresis assay, we identify replication origins (ORI7-LI and ORI7-RI) proximal to an early replicating centromere (CEN7) in C. albicans. We show that the replication forks stall at CEN7 in a kinetochore dependent manner and fork stalling is reduced in the absence of the homologous recombination (HR) proteins Rad51 and Rad52. Deletion of ORI7-RI causes a significant reduction in the stalled fork signal and an increased loss rate of the altered chromosome 7. The HR proteins, Rad51 and Rad52, have been shown to play a role in fork restart. Confocal microscopy shows declustered kinetochores in rad51 and rad52 mutants, which are evidence of kinetochore disintegrity. CENP-ACaCse4 levels at centromeres, as determined by chromatin immunoprecipitation (ChIP) experiments, are reduced in absence of Rad51/Rad52 resulting in disruption of the kinetochore structure. Moreover, western blot analysis reveals that delocalized CENP-A molecules in HR mutants degrade in a similar fashion as in other kinetochore mutants described before. Finally, co-immunoprecipitation assays indicate that Rad51 and Rad52 physically interact with CENP-ACaCse4 in vivo. Thus, the HR proteins Rad51 and Rad52 epigenetically maintain centromere functioning by regulating CENP-ACaCse4 levels at the programmed stall sites of early replicating centromeres.
在大多数真核生物中,着丝粒位置的确定并非仅取决于DNA序列。然而,着丝粒身份的非遗传决定因素尚未明确界定。虽然多种机制单独或协同作用可能在表观遗传上确定着丝粒,但该领域的大多数研究都集中在一个普遍因素上,即着丝粒特异性组蛋白H3变体CENP-A,它常被视为着丝粒身份的表观遗传决定因素。尽管跨物种在着丝粒处加载CENP-A的时间各不相同,但在大多数酵母着丝粒中发现了与复制相关的CENP-A在S期早期沉积。着丝粒是致病性芽殖酵母白色念珠菌中最早复制的染色体区域。使用二维琼脂糖凝胶电泳分析,我们在白色念珠菌中鉴定出靠近早期复制着丝粒(CEN7)的复制起点(ORI7-LI和ORI7-RI)。我们发现复制叉以依赖动粒的方式在CEN7处停滞,并且在没有同源重组(HR)蛋白Rad51和Rad52的情况下,叉停滞减少。删除ORI7-RI会导致停滞叉信号显著降低,并且改变的7号染色体的丢失率增加。HR蛋白Rad51和Rad52已被证明在叉重新启动中起作用。共聚焦显微镜显示rad51和rad52突变体中的动粒分散,这是动粒完整性破坏的证据。通过染色质免疫沉淀(ChIP)实验确定,在没有Rad51/Rad52的情况下,着丝粒处的CENP-ACaCse4水平降低,导致动粒结构破坏。此外,蛋白质印迹分析表明,HR突变体中异位的CENP-A分子与之前描述的其他动粒突变体中的降解方式相似。最后,免疫共沉淀分析表明,Rad51和Rad52在体内与CENP-ACaCse4发生物理相互作用。因此,HR蛋白Rad51和Rad52通过调节早期复制着丝粒的程序性停滞位点处的CENP-ACaCse4水平,在表观遗传上维持着丝粒的功能。