Duneau David, Lafont Pierre D M, Lauzeral Christine, Parthuisot Nathalie, Faucher Christian, Jin Xuerong, Buchon Nicolas, Ferdy Jean-Baptiste
Centre de Recherche sur la Biodiversit´e et l'Environnement, Universit´e Paul Sabatier, Toulouse, France.
Centre for Cardiovascular Sciences, Queen's Medical Research Institute, University of Edinburgh, Edinburgh, United Kingdom.
Elife. 2025 Feb 13;14:e104052. doi: 10.7554/eLife.104052.
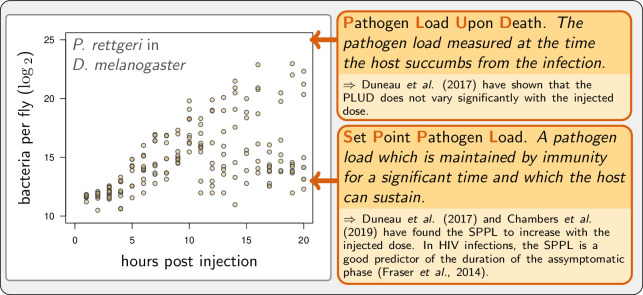
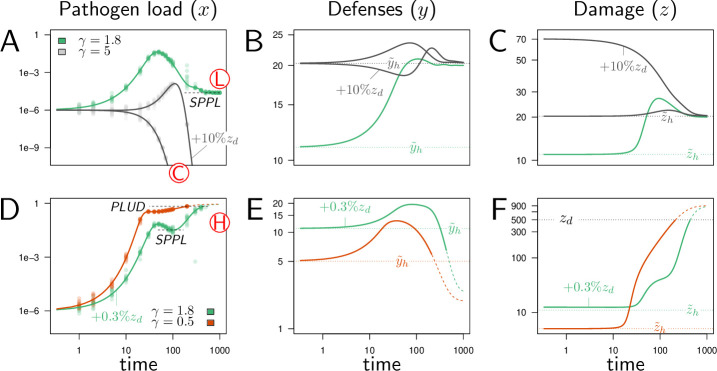
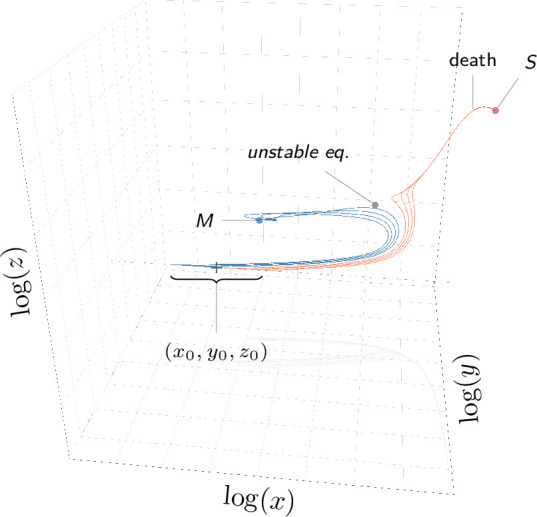
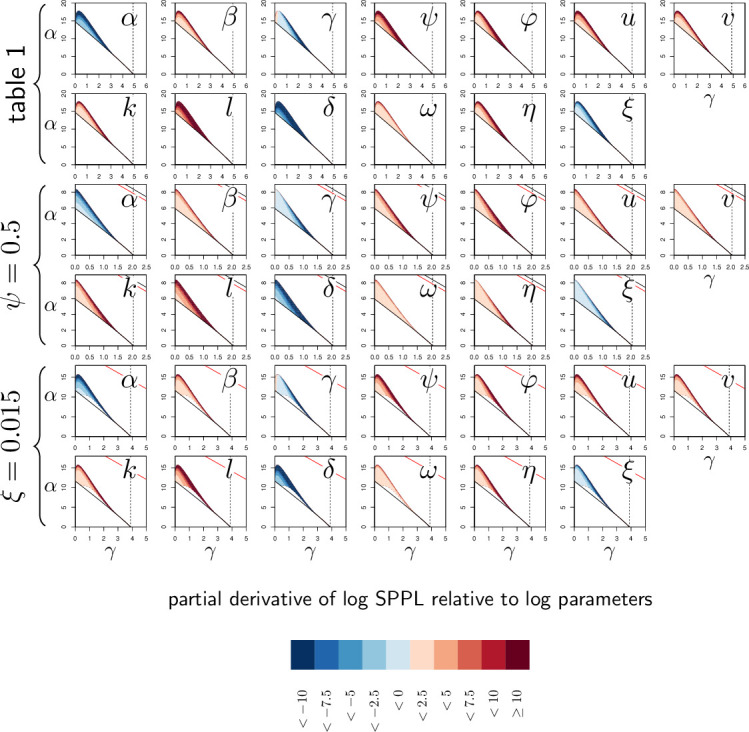
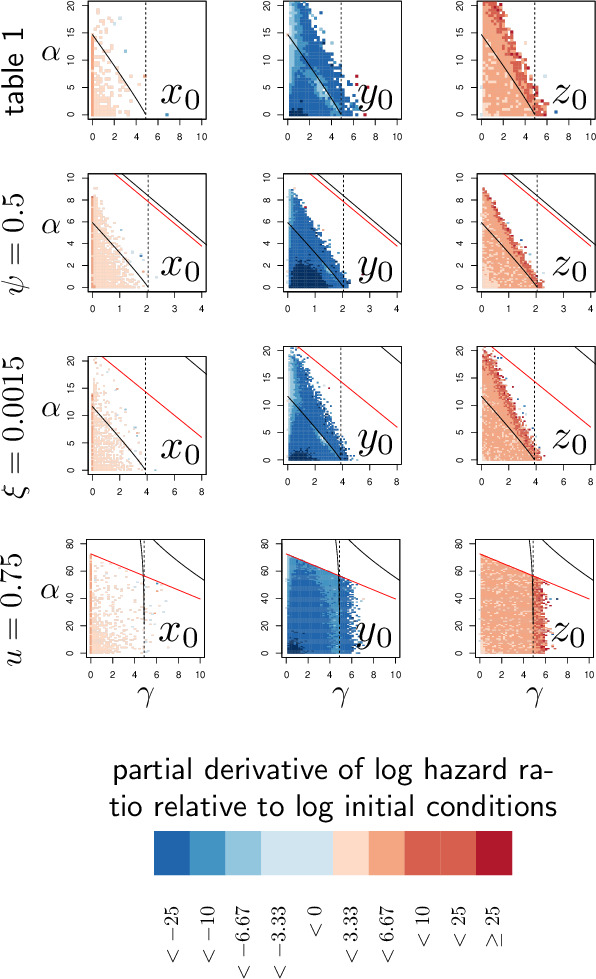
How are some individuals surviving infections while others die? The answer lies in how infected individuals invest into controlling pathogen proliferation and mitigating damage, two strategies respectively called resistance and disease tolerance. Pathogen within-host dynamics (WHD), influenced by resistance, and its connection to host survival, determined by tolerance, decide the infection outcome. To grasp these intricate effects of resistance and tolerance, we used a deterministic theoretical model where pathogens interact with the immune system of a host. The model describes the positive and negative regulation of the immune response, consider the way damage accumulate during the infection and predicts WHD. When chronic, infections stabilize at a Set-Point Pathogen Load (SPPL). Our model predicts that this situation can be transient, the SPPL being then a predictor of life span which depends on initial condition (e.g. inoculum). When stable, the SPPL is rather diagnostic of non-lethal chronic infections. In lethal infections, hosts die at a Pathogen Load Upon Death (PLUD) which is almost independent from the initial conditions. As the SPPL, the PLUD is affected by both resistance and tolerance but we demonstrate that it can be used in conjunction with mortality measurement to distinguish the effect of disease tolerance from that of resistance. We validate empirically this new approach, using and the pathogen . We found that, as predicted by the model, hosts that were wounded or deficient of key antimicrobial peptides had a higher PLUD, while Catalase mutant hosts, likely to have a default in disease tolerance, had a lower PLUD.
为什么有些人在感染中存活下来而另一些人却死亡?答案在于受感染个体如何投入资源来控制病原体增殖和减轻损害,这两种策略分别称为抗性和耐病性。受抗性影响的病原体宿主体内动态(WHD)及其与由耐病性决定的宿主存活之间的联系,决定了感染的结果。为了理解抗性和耐病性的这些复杂影响,我们使用了一个确定性理论模型,其中病原体与宿主的免疫系统相互作用。该模型描述了免疫反应的正负调节,考虑了感染过程中损害积累的方式并预测WHD。当感染为慢性时,感染会在设定点病原体负荷(SPPL)处稳定下来。我们的模型预测这种情况可能是短暂的,此时SPPL是寿命的一个预测指标,它取决于初始条件(例如接种量)。当处于稳定状态时,SPPL更能诊断非致命性慢性感染。在致命性感染中,宿主在死亡时的病原体负荷(PLUD)下死亡,而PLUD几乎与初始条件无关。与SPPL一样,PLUD受抗性和耐病性两者的影响,但我们证明它可以与死亡率测量结合使用,以区分耐病性和抗性的影响。我们使用 和病原体通过实验验证了这种新方法。我们发现,正如模型所预测的那样,受伤或缺乏关键抗菌肽的宿主具有更高的PLUD,而过氧化氢酶突变体宿主可能在耐病性方面存在缺陷,其PLUD较低。