Nguyen Mai, Bauda Elda, Boyat Célia, Laguri Cédric, Freton Céline, Chouquet Anne, Gallet Benoit, Baudoin Morgane, Wong Yung-Sing, Grangeasse Christophe, Moriscot Christine, Durmort Claire, Zapun André, Morlot Cecile
Univ. Grenoble Alpes, CNRS, CEA, IBS, Grenoble, France.
Univ. Grenoble Alpes, CNRS, DPM, Grenoble, France.
Elife. 2025 Apr 23;14:RP105132. doi: 10.7554/eLife.105132.
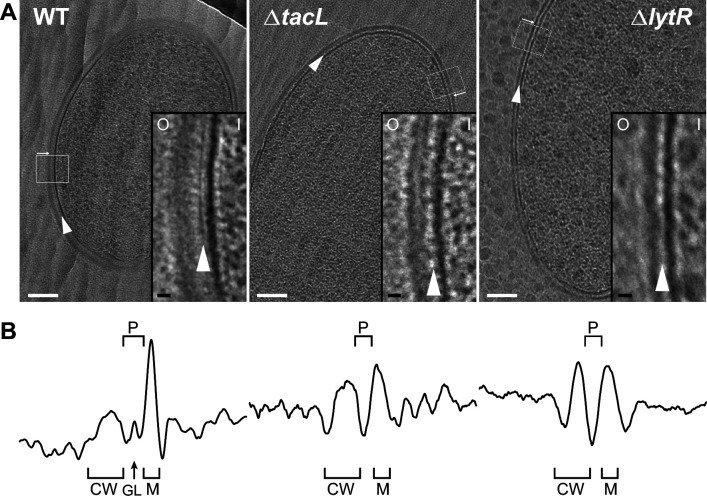
Teichoic acids (TA) are linear phospho-saccharidic polymers and important constituents of the cell envelope of Gram-positive bacteria, either bound to the peptidoglycan as wall teichoic acids (WTA) or to the membrane as lipoteichoic acids (LTA). The composition of TA varies greatly but the presence of both WTA and LTA is highly conserved, hinting at an underlying fundamental function that is distinct from their specific roles in diverse organisms. We report the observation of a periplasmic space in by cryo-electron microscopy of vitreous sections. The thickness and appearance of this region change upon deletion of genes involved in the attachment of TA, supporting their role in the maintenance of a periplasmic space in Gram-positive bacteria as a possible universal function. Consequences of these mutations were further examined by super-resolved microscopy, following metabolic labeling and fluorophore coupling by click chemistry. This novel labeling method also enabled in-gel analysis of cell fractions. With this approach, we were able to titrate the actual amount of TA per cell and to determine the ratio of WTA to LTA. In addition, we followed the change of TA length during growth phases, and discovered that a mutant devoid of LTA accumulates the membrane-bound polymerized TA precursor.
磷壁酸(TA)是线性磷酸化糖聚合物,是革兰氏阳性菌细胞壁的重要组成成分,可作为壁磷壁酸(WTA)与肽聚糖结合,或作为脂磷壁酸(LTA)与细胞膜结合。TA的组成差异很大,但WTA和LTA的存在高度保守,这暗示着其具有不同于它们在不同生物体中特定作用的潜在基本功能。我们通过对玻璃切片进行冷冻电子显微镜观察,发现了[具体研究对象]中的周质空间。该区域的厚度和外观在参与TA附着的基因缺失后发生变化,这支持了它们在维持革兰氏阳性菌周质空间中可能具有普遍功能的作用。通过超分辨显微镜,在代谢标记和点击化学荧光团偶联后,进一步研究了这些突变的后果。这种新颖的标记方法还能够对细胞组分进行凝胶内分析。通过这种方法,我们能够滴定每个细胞中TA的实际含量,并确定WTA与LTA的比例。此外,我们跟踪了生长阶段TA长度的变化,发现缺乏LTA的突变体积累了膜结合的聚合TA前体。