Grootwassink J W, Gaucher G M
J Bacteriol. 1980 Feb;141(2):443-55. doi: 10.1128/jb.141.2.443-455.1980.
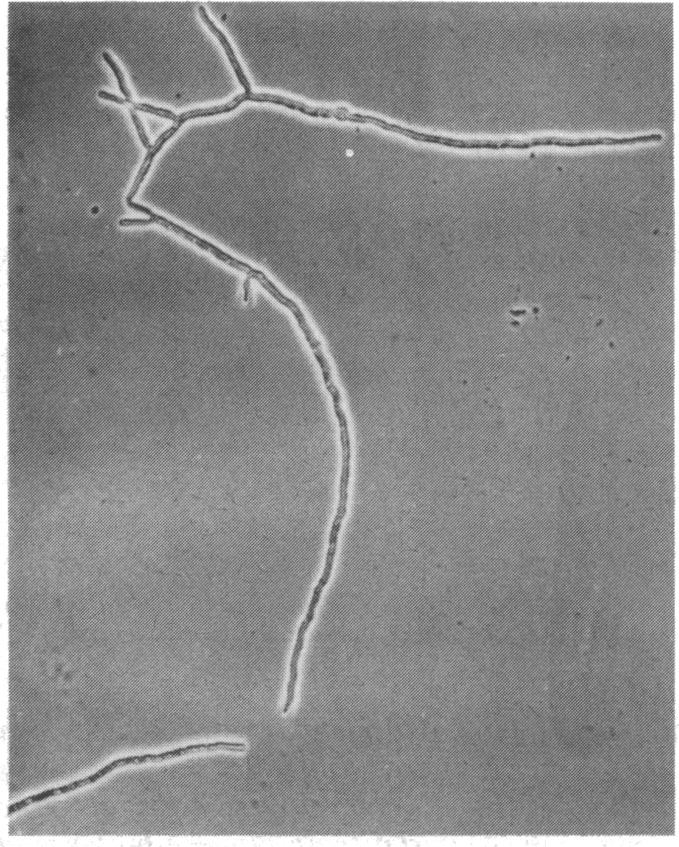
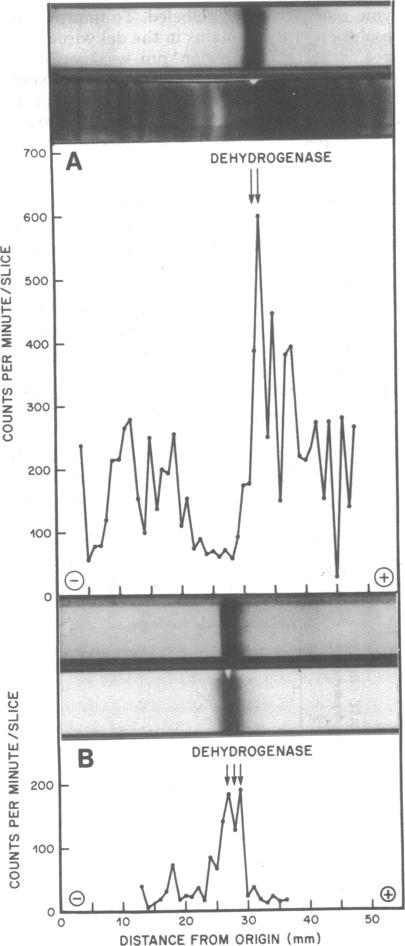
The initiation of patulin biosynthesis in submerged batch cultures of Penicillium urticae NRRL 2159A was investigated at the enzyme level. In contrast to earlier studies, this study achieved a clear temporal separation of growing cells devoid of secondary metabolism-specific enzymes from nongrowing cells, which rapidly produce these enzymes. A spore inoculum, silicone-treated flasks, and two new media which supported a rapid, pellet-free, filamentous type of growth were used. In yeast extract-glucose-buffer medium, a marked drop in the specific growth rate (approximately equal to 0.26 h-1) coincided with the appearance of the first pathway-specific enzyme, 6-methylsalicylic acid synthetase, at about 19 h after inoculation. About 3 h later, when replicatory growth had ceased entirely, the sparsely branched mycelia (length, approximately equal to 550 microns) began the rapid synthesis of a later pathway enzyme, m-hydroxybenzyl alcohol dehydrogenase. A similar sequence of events occurred in a defined nitrate-glucose-buffer medium; 12 other strains or isolates of P. urticae, as well as some patulin-producing aspergilli, behaved in a similar manner. The age at which a culture produced m-hydroxybenzyl alcohol dehydrogenase was increased by increasing the nutrient nitrogen content of the medium or by decreasing the size of the spore inoculum. In each instance the appearance of enzyme was determined by the nutritional status of the culture and not by its age. A similar appearance of patulin pathway enzymes occurred when a growing culture was resuspended in a nitrogen-free 4% glucose solution with or without 0.1 M phosphate (pH 6.5). The appearance of both the synthetase and the dehydrogenase was arrested by the addition of cycloheximide (0.4 to 5 micrograms/ml) or actinomycin D (20 to 80 micrograms/ml). This requirement for de novo protein and ribonucleic acid syntheses was confirmed by the incorporation of labeled leucine into the dehydrogenase, and the possibility that latent or preformed proteins were being activated was eliminated.
在酶水平上研究了荨麻青霉NRRL 2159A深层分批培养中棒曲霉素生物合成的起始。与早期研究不同,本研究实现了无次生代谢特异性酶的生长细胞与快速产生这些酶的非生长细胞在时间上的清晰分离。使用了孢子接种物、经硅酮处理的烧瓶以及两种支持快速、无颗粒、丝状生长类型的新培养基。在酵母提取物 - 葡萄糖 - 缓冲液培养基中,比生长速率显著下降(约等于0.26 h⁻¹)与接种后约19 h时第一种途径特异性酶6 - 甲基水杨酸合成酶的出现同时发生。大约3 h后,当复制性生长完全停止时,稀疏分支的菌丝体(长度约等于550微米)开始快速合成后期途径酶间羟基苄醇脱氢酶。在限定的硝酸盐 - 葡萄糖 - 缓冲液培养基中发生了类似的一系列事件;荨麻青霉的其他12个菌株或分离株以及一些产棒曲霉素的曲霉也表现出类似的行为。通过增加培养基中的营养氮含量或减小孢子接种物的大小,培养物产生间羟基苄醇脱氢酶的年龄会增加。在每种情况下,酶的出现取决于培养物的营养状态而非其年龄。当生长的培养物重悬于含或不含0.1 M磷酸盐(pH 6.5)的无氮4%葡萄糖溶液中时,棒曲霉素途径酶出现了类似情况。合成酶和脱氢酶的出现都因添加放线菌酮(0.4至5微克/毫升)或放线菌素D(20至80微克/毫升)而停止。通过将标记的亮氨酸掺入脱氢酶中证实了对从头蛋白质和核糖核酸合成的这种需求,并排除了潜在或预先形成的蛋白质被激活的可能性。