Bost S, Belin D
Départment de Pathologie, Université de Genève, Switzerland.
EMBO J. 1995 Sep 15;14(18):4412-21. doi: 10.1002/j.1460-2075.1995.tb00120.x.
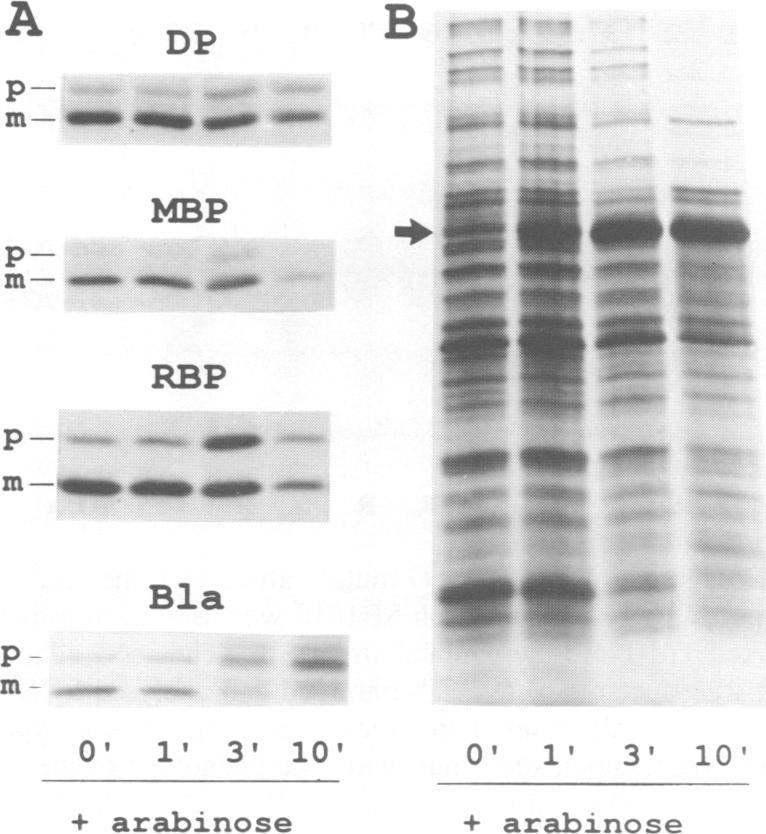
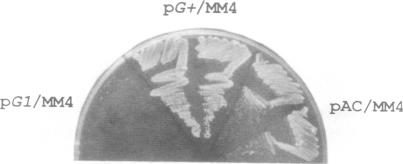
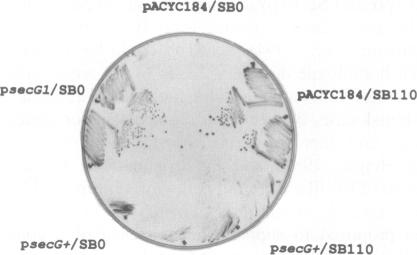
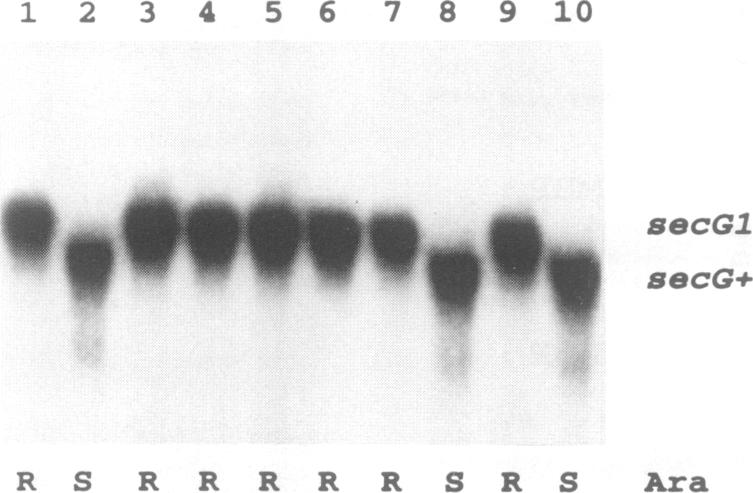
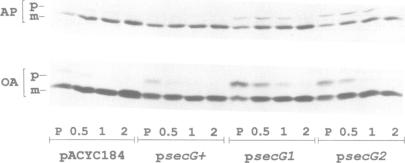
The signal sequence of the murine serine protease inhibitor PAI-2 promotes alkaline phosphatase export to the E. coli periplasm. However, high level expression of this chimeric protein interferes with cell growth. Since most suppressors of this toxic phenotype map to secA and secY, growth arrest results from a defective interaction of the chimeric protein with the export machinery. We have characterized suppressors which map in secG, a newly defined gene of the export machinery. All single amino acid substitutions map to three adjacent codons. These secG mutants have a weak Sec phenotype, as determined by their effect on export mediated by wild-type and mutant signal sequences. Whilst a secG disruption allele also confers a weak Sec phenotype, it does not suppress the toxicity of the chimeric protein. This difference results from a selective effect of the secG suppressors on the kinetics of export mediated by the PAI-2 signal sequence. Using a malE signal sequence mutant, which has a Mal-phenotype in secG mutant strains, we have isolated extragenic Mal+ suppressors. Most suppressors map to secY, and several are allele-specific. Finally, SecG overexpression accelerates the kinetics of protein export, suggesting that there are two types of functional translocation complexes: with or without SecG.
小鼠丝氨酸蛋白酶抑制剂PAI-2的信号序列可促进碱性磷酸酶输出至大肠杆菌周质。然而,这种嵌合蛋白的高水平表达会干扰细胞生长。由于这种毒性表型的大多数抑制子定位于secA和secY,生长停滞是由嵌合蛋白与输出机制的缺陷性相互作用导致的。我们已经鉴定了定位于secG的抑制子,secG是输出机制中的一个新定义基因。所有单氨基酸替换都定位于三个相邻密码子。这些secG突变体具有较弱的Sec表型,这是通过它们对野生型和突变信号序列介导的输出的影响来确定的。虽然secG缺失等位基因也赋予较弱的Sec表型,但它不能抑制嵌合蛋白的毒性。这种差异是由secG抑制子对PAI-2信号序列介导的输出动力学的选择性作用导致的。使用在secG突变菌株中具有Mal表型的malE信号序列突变体,我们分离出了基因外的Mal+抑制子。大多数抑制子定位于secY,并且有几个是等位基因特异性的。最后,SecG的过表达加速了蛋白质输出的动力学,表明存在两种类型的功能性转运复合物:有或没有SecG。