Dieci G, Hermann-Le Denmat S, Lukhtanov E, Thuriaux P, Werner M, Sentenac A
Service de Biochimie et Génétique Moléculaire, CEA-Saclay, Gif-sur-Yvette, France.
EMBO J. 1995 Aug 1;14(15):3766-76. doi: 10.1002/j.1460-2075.1995.tb00046.x.
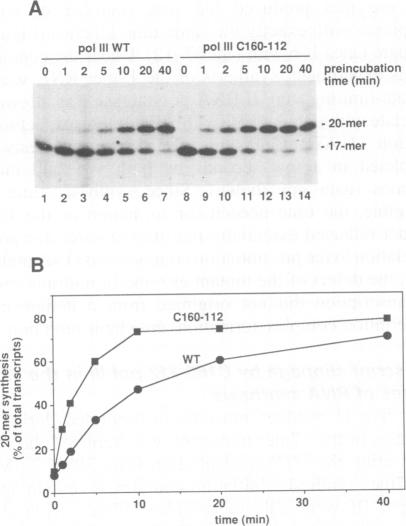
The largest subunits of the three eukaryotic nuclear RNA polymerase present extensive sequence homology with the beta' subunit of the bacterial enzymes over five major co-linear regions. Region d is the most highly conserved and contains a motif, (Y/F)NADFDGD(E/Q)M(N/A), which is invariant in all multimeric RNA polymerases. An extensive mutagenesis of that region in yeast RNA polymerase III led to a vast majority (16/22) of lethal single-site substitutions. A few conditional mutations were also obtained. One of them, rpc160-112, corresponds to a double substitution (T506I, N509Y) and has a slow growth phenotype at 25 degrees C. RNA polymerase III from the mutant rpc160-112 was severely impaired in its ability to transcribe a tRNA gene in vitro. The transcription defect did not originate from a deficiency in transcription complex formation and RNA chain initiation, but was mainly due to a reduced elongation rate. Under conditions of substrate limitation, the mutant enzyme showed increased pausing at the intrinsic pause sites of the SUP4 tRNA gene and an increased rate of slippage of nascent RNA, as compared with the wild-type enzyme. The enzyme defect was also detectable with poly[d(A-T)] as template, in the presence of saturating DNA, ATP and UTP concentrations. The mutant enzyme behavior is best explained by a distortion of the active site near the growing point of the RNA product.
三种真核细胞核RNA聚合酶的最大亚基在五个主要共线区域与细菌酶的β'亚基存在广泛的序列同源性。区域d是最保守的,包含一个基序(Y/F)NADFDGD(E/Q)M(N/A),在所有多聚体RNA聚合酶中都是不变的。对酵母RNA聚合酶III中该区域进行广泛的诱变导致绝大多数(16/22)致死性单点取代。还获得了一些条件突变。其中一个rpc160 - 112对应于双取代(T506I, N509Y),在25℃时具有生长缓慢的表型。来自突变体rpc160 - 112的RNA聚合酶III在体外转录tRNA基因的能力严重受损。转录缺陷并非源于转录复合物形成和RNA链起始的缺陷,而是主要由于延伸率降低。在底物限制条件下,与野生型酶相比,突变酶在SUP4 tRNA基因的内在暂停位点处的暂停增加,新生RNA的滑动速率增加。在饱和DNA、ATP和UTP浓度存在的情况下,以聚[d(A - T)]为模板也可检测到该酶缺陷。突变酶的行为最好用RNA产物生长点附近活性位点的扭曲来解释。