Sturchler C, Lescure A, Keith G, Carbon P, Krol A
UPR du CNRS Structure des Macromolécules Biologiques et Mécanismes de Reconnaissance, Strasbourg, France.
Nucleic Acids Res. 1994 Apr 25;22(8):1354-8. doi: 10.1093/nar/22.8.1354.
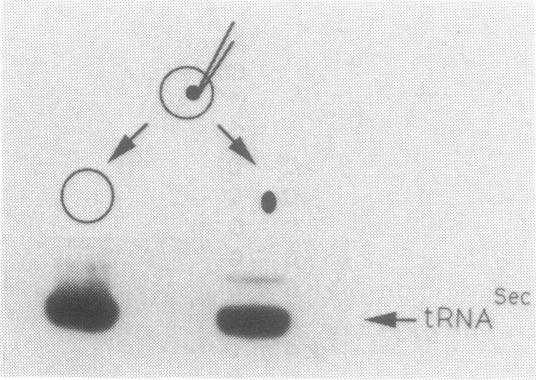
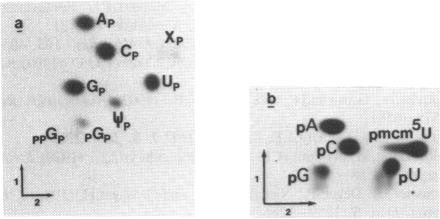
We examined the base modification pattern of Xenopus tRNA(Sec) using microinjection into Xenopus oocytes, with particular focus on the wobble base U34 at the first position of the anticodon. We found that U34 becomes modified to mcm5U34 (5-methylcarboxymethyluridine) in the oocyte cytoplasm in a rather complex manner. When the tRNA(Sec) gene is injected into Xenopus oocyte nuclei, psi 55 and m1A58 are readily obtained, but not mcm5U34. This will appear only upon cytoplasmic injection of the gene product arising from the first nuclear injection. In contrast, tRNA(Sec) produced by in vitro transcription with T7 RNA polymerase readily acquires i6A37, psi 55, m1A58, and mcm5U34. The latter is obtained after direct nuclear or cytoplasmic injections. It has been reported by others that mcm5Um, a 2'-O-methylated derivative of mcm5U34, also exists in rat and bovine tRNA(Sec). With both the gene product and the in vitro transcript, and using the sensitive RNase T2 assay, we were unable to detect under our conditions the presence of a dinucleotide carrying mcm5Um and that would be therefore refractory to hydrolysis. We showed that the unusual mcm5U acquisition pathway does not result from impairment of nucleocytoplasmic transport. Rather, these data can be interpreted to mean that the modification is performed by a tRNA(Sec) specific enzyme, limiting in the oocyte cytoplasm.
我们通过显微注射到非洲爪蟾卵母细胞中,研究了非洲爪蟾tRNA(Sec)的碱基修饰模式,特别关注反密码子第一位的摆动碱基U34。我们发现U34在卵母细胞细胞质中以相当复杂的方式被修饰为mcm5U34(5-甲基羧甲基尿苷)。当将tRNA(Sec)基因注射到非洲爪蟾卵母细胞核中时,很容易获得ψ55和m1A58,但没有mcm5U34。只有在将第一次核注射产生的基因产物进行细胞质注射后才会出现mcm5U34。相比之下,用T7 RNA聚合酶体外转录产生的tRNA(Sec)很容易获得i6A37、ψ55、m1A58和mcm5U34。后者在直接进行核注射或细胞质注射后即可获得。其他人曾报道,mcm5U34的2'-O-甲基化衍生物mcm5Um也存在于大鼠和牛的tRNA(Sec)中。对于基因产物和体外转录本,使用灵敏的核糖核酸酶T2检测法,在我们的实验条件下,我们无法检测到携带mcm5Um的二核苷酸的存在及其对水解的抗性。我们表明,这种不寻常的mcm5U获得途径并非由核质运输受损导致。相反,这些数据可以解释为这种修饰是由一种tRNA(Sec)特异性酶进行的,该酶在卵母细胞细胞质中受到限制。