Klenova E M, Nicolas R H, Paterson H F, Carne A F, Heath C M, Goodwin G H, Neiman P E, Lobanenkov V V
Institute of Carcinogenesis, Russian Cancer Research Center, Moscow.
Mol Cell Biol. 1993 Dec;13(12):7612-24. doi: 10.1128/mcb.13.12.7612-7624.1993.
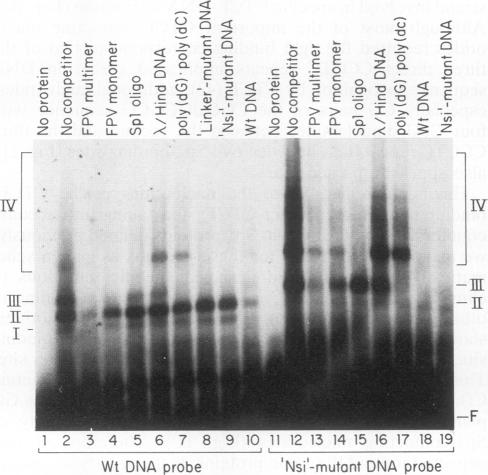
A novel sequence-specific DNA-binding protein, CTCF, which interacts with the chicken c-myc gene promoter, has been identified and partially characterized (V. V. Lobanenkov, R. H. Nicolas, V. V. Adler, H. Paterson, E. M. Klenova, A. V. Polotskaja, and G. H. Goodwin, Oncogene 5:1743-1753, 1990). In order to test directly whether binding of CTCF to one specific DNA region of the c-myc promoter is important for chicken c-myc transcription, we have determined which nucleotides within this GC-rich region are responsible for recognition of overlapping sites by CTCF and Sp1-like proteins. Using missing-contact analysis of all four nucleotides in both DNA strands and homogeneous CTCF protein purified by sequence-specific chromatography, we have identified three sets of nucleotides which contact either CTCF or two Sp1-like proteins binding within the same DNA region. Specific mutations of 3 of 15 purines required for CTCF binding were designed to eliminate binding of CTCF without altering the binding of other proteins. Electrophoretic mobility shift assay of nuclear extracts showed that the mutant DNA sequence did not bind CTCF but did bind two Sp1-like proteins. When introduced into a 3.3-kbp-long 5'-flanking noncoding c-myc sequence fused to a reporter CAT gene, the same mutation of the CTCF binding site resulted in 10- and 3-fold reductions, respectively, of transcription in two different (erythroid and myeloid) stably transfected chicken cell lines. Isolation and analysis of the CTCF cDNA encoding an 82-kDa form of CTCF protein shows that DNA-binding domain of CTCF is composed of 11 Zn fingers: 10 are of C2H2 class, and 1 is of C2HC class. CTCF was found to be abundant and conserved in cells of vertebrate species. We detected six major nuclear forms of CTCF protein differentially expressed in different chicken cell lines and tissues. We conclude that isoforms of 11-Zn-finger factor CTCF which are present in chicken hematopoietic HD3 and BM2 cells can act as a positive regulator of the chicken c-myc gene transcription. Possible functions of other CTCF forms are discussed.
一种与鸡c-myc基因启动子相互作用的新型序列特异性DNA结合蛋白CTCF已被鉴定并部分表征(V.V.洛巴年科夫、R.H.尼古拉斯、V.V.阿德勒、H.帕特森、E.M.克列诺娃、A.V.波洛茨卡娅和G.H.古德温,《癌基因》5:1743 - 1753,1990)。为了直接测试CTCF与c-myc启动子的一个特定DNA区域的结合对鸡c-myc转录是否重要,我们确定了这个富含GC的区域内哪些核苷酸负责CTCF和Sp1样蛋白对重叠位点的识别。通过对两条DNA链上所有四个核苷酸进行缺失接触分析以及使用序列特异性色谱法纯化的均一CTCF蛋白,我们确定了三组与CTCF或在同一DNA区域内结合的两种Sp1样蛋白接触的核苷酸。设计了CTCF结合所需的15个嘌呤中的3个的特异性突变,以消除CTCF的结合而不改变其他蛋白质的结合。核提取物的电泳迁移率变动分析表明,突变的DNA序列不结合CTCF,但结合两种Sp1样蛋白。当将相同的CTCF结合位点突变引入与报告基因CAT基因融合的3.3 kbp长的5'侧翼非编码c-myc序列中时,在两种不同的(红细胞系和髓细胞系)稳定转染的鸡细胞系中,转录分别降低了10倍和3倍。对编码82 kDa形式的CTCF蛋白的CTCF cDNA进行分离和分析表明,CTCF的DNA结合结构域由11个锌指组成:10个是C2H2类,1个是C2HC类。发现CTCF在脊椎动物物种的细胞中含量丰富且保守。我们在不同的鸡细胞系和组织中检测到六种主要的CTCF蛋白核形式差异表达。我们得出结论,存在于鸡造血HD3和BM2细胞中的11锌指因子CTCF的同工型可以作为鸡c-myc基因转录的正调节因子。还讨论了其他CTCF形式的可能功能。