Simmons D T, Upson R, Wun-Kim K, Young W
School of Life and Health Sciences, University of Delaware, Newark 19716.
J Virol. 1993 Jul;67(7):4227-36. doi: 10.1128/JVI.67.7.4227-4236.1993.
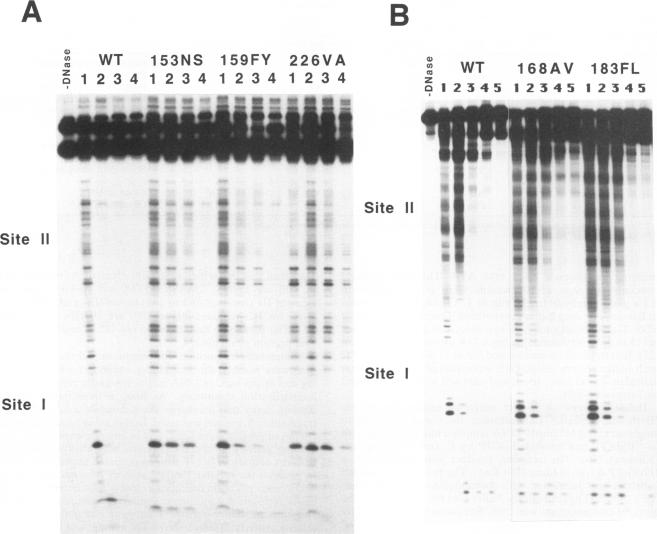
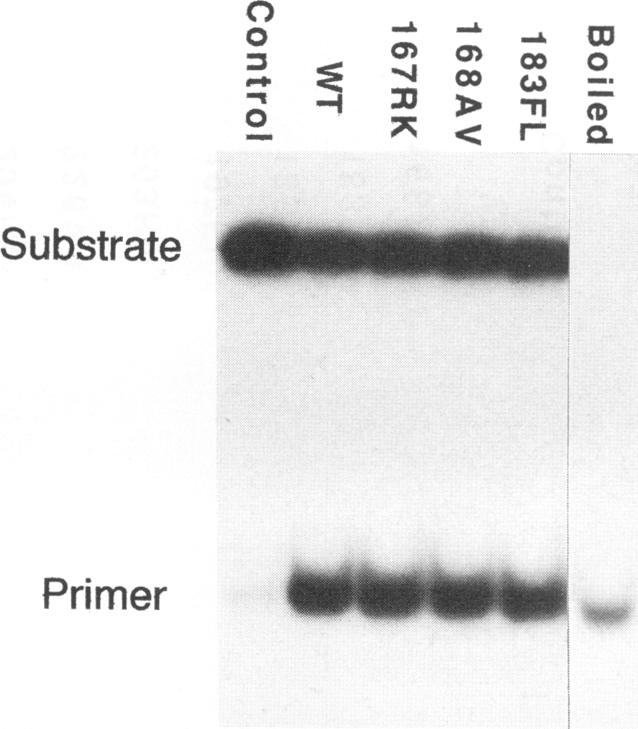
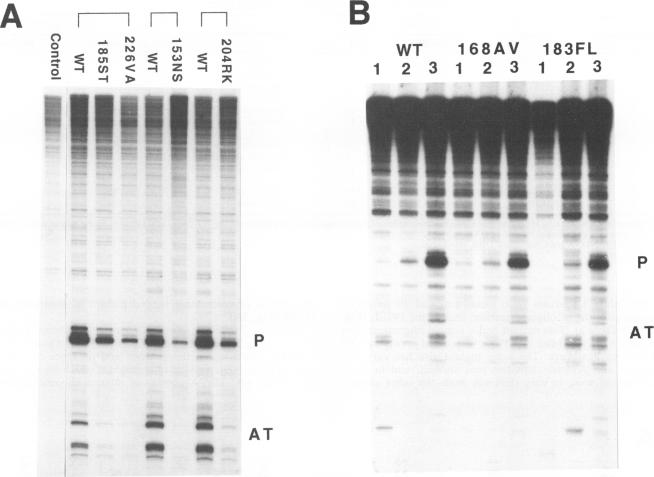
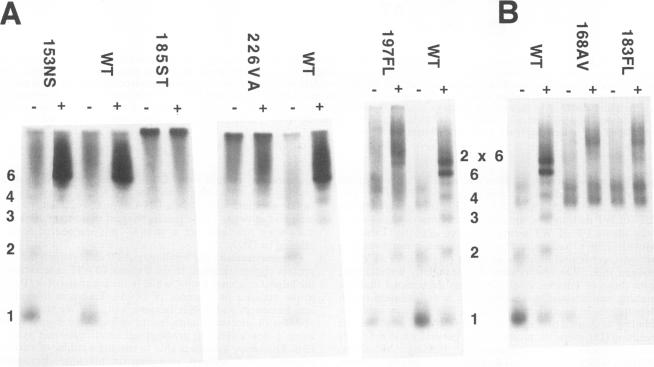
The role of the origin-binding domain of simian virus 40 large tumor antigen (T antigen) in the initiation of virus DNA replication was investigated by analyzing the biochemical activities of a series of mutants with single-site substitutions in this region. These activities include origin-specific and nonspecific DNA binding, melting of the imperfect palindromic sequence, untwisting of the AT-rich region, unwinding of origin-containing DNA, helicase activity, and the ability to oligomerize normally in response to ATP. Three classes of T-antigen mutants that are unable to support virus replication in monkey cells are described. Class 1 mutants are unable to bind to the origin of DNA replication but are able to bind to DNA nonspecifically. Class 2 mutants exhibit defective binding to both types of DNA. As expected, mutants in these first two classes are unable to unwind origin DNA. Surprisingly, however, these mutants possess significant levels of melting and untwisting activities, suggesting that these reactions may not be solely dependent on the ability of the protein to recognize origin sequences. Most class 1 mutants oligomerize normally in response to ATP, indicating that their DNA-binding defects are not due to structural alterations but probably to a failure to directly recognize origin sequences. In contrast, class 2 mutants exhibit defective oligomerization. Class 3 mutants bind to origin and nonorigin DNA at near wild-type levels and melt and untwist origin DNA normally but exhibit defective oligomerization and unwinding. These mutants are, however, perfectly able to carry out the helicase reaction, indicating that their unwinding defect is at some step after melting but before a nonspecific helicase is used to separate parental strands during replication. These results therefore suggest that proper oligomerization to correctly position the molecules on the DNA may be more important in initiating unwinding than in bringing about efficient DNA binding, inducing structural changes in the DNA, or carrying out the helicase reaction.
通过分析该区域一系列单点取代突变体的生化活性,研究了猿猴病毒40大T抗原(T抗原)的起始结合结构域在病毒DNA复制起始中的作用。这些活性包括起始位点特异性和非特异性DNA结合、不完全回文序列的解链、富含AT区域的解旋、含起始位点DNA的解旋、解旋酶活性以及响应ATP正常寡聚化的能力。描述了三类无法在猴细胞中支持病毒复制的T抗原突变体。1类突变体无法结合DNA复制起始位点,但能够非特异性地结合DNA。2类突变体对两种类型的DNA结合均有缺陷。正如预期的那样,前两类突变体无法解旋起始位点DNA。然而,令人惊讶的是,这些突变体具有显著水平的解链和解旋活性,这表明这些反应可能不仅仅取决于蛋白质识别起始序列的能力。大多数1类突变体响应ATP正常寡聚化,表明它们的DNA结合缺陷不是由于结构改变,而可能是由于未能直接识别起始序列。相比之下,2类突变体表现出寡聚化缺陷。3类突变体以接近野生型的水平结合起始位点和非起始位点DNA,正常解链和解旋起始位点DNA,但表现出寡聚化和解旋缺陷。然而,这些突变体完全能够进行解旋酶反应,表明它们的解旋缺陷发生在解链后的某个步骤,但在复制过程中使用非特异性解旋酶分离亲本链之前。因此,这些结果表明,在起始解链过程中,正确寡聚化以使分子在DNA上正确定位可能比实现有效的DNA结合、诱导DNA结构变化或进行解旋酶反应更为重要。