Xu Z, Mayer A, Muller E, Wickner W
Department of Biochemistry, Dartmouth Medical School, Hanover, New Hampshire 03755-3844, USA.
J Cell Biol. 1997 Jan 27;136(2):299-306. doi: 10.1083/jcb.136.2.299.
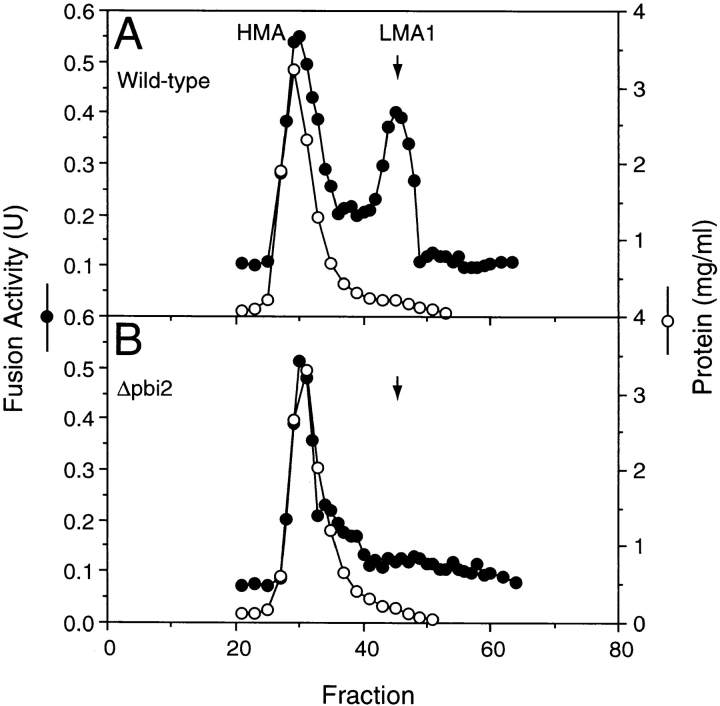
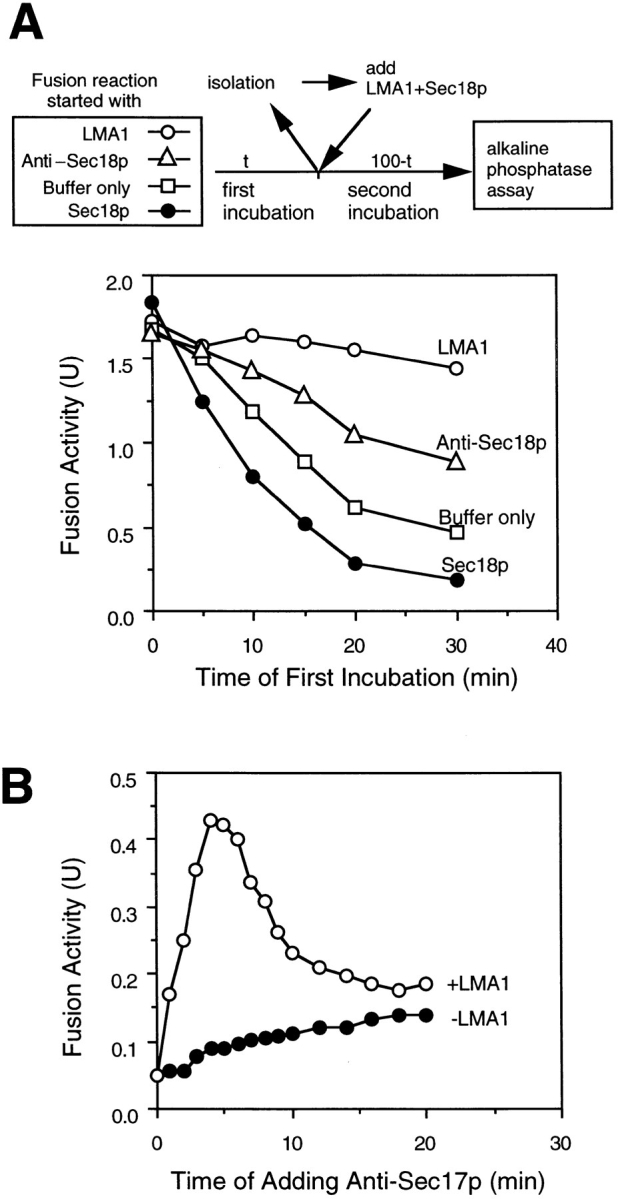
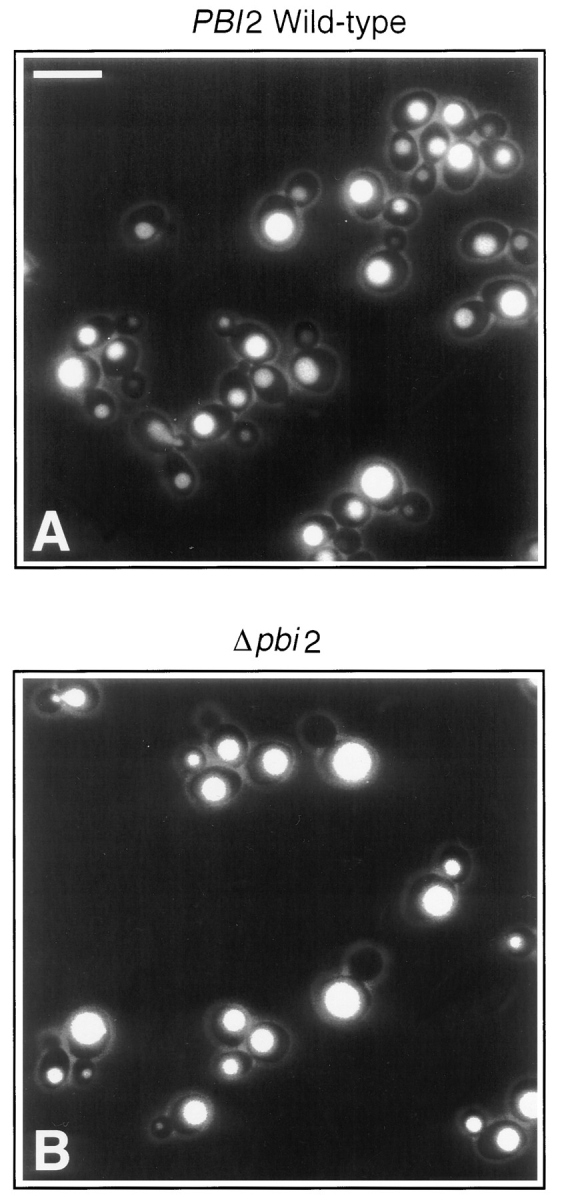
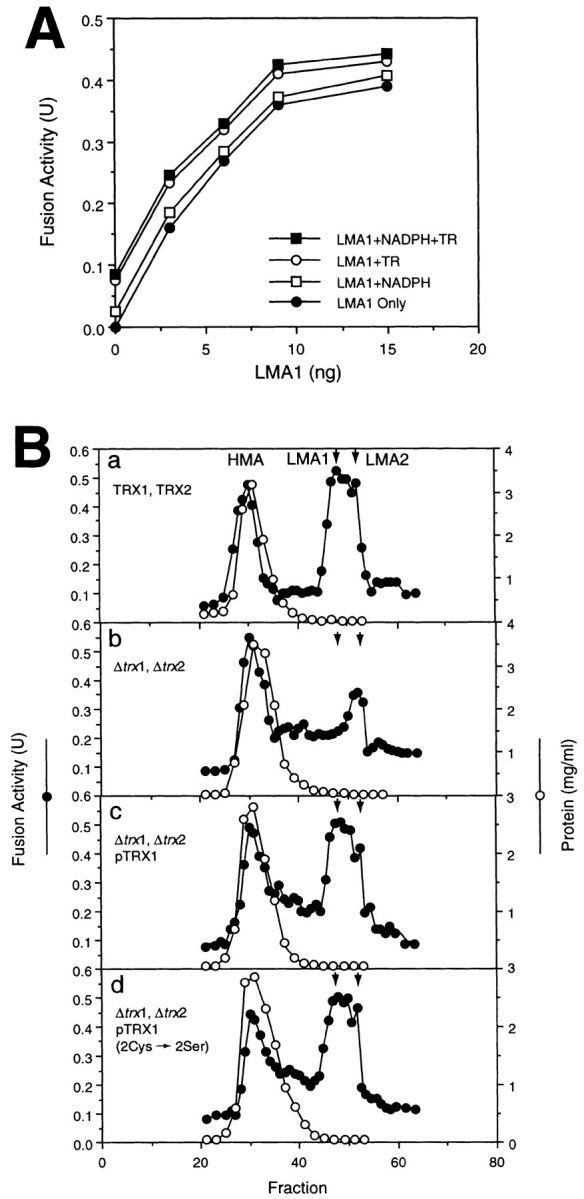
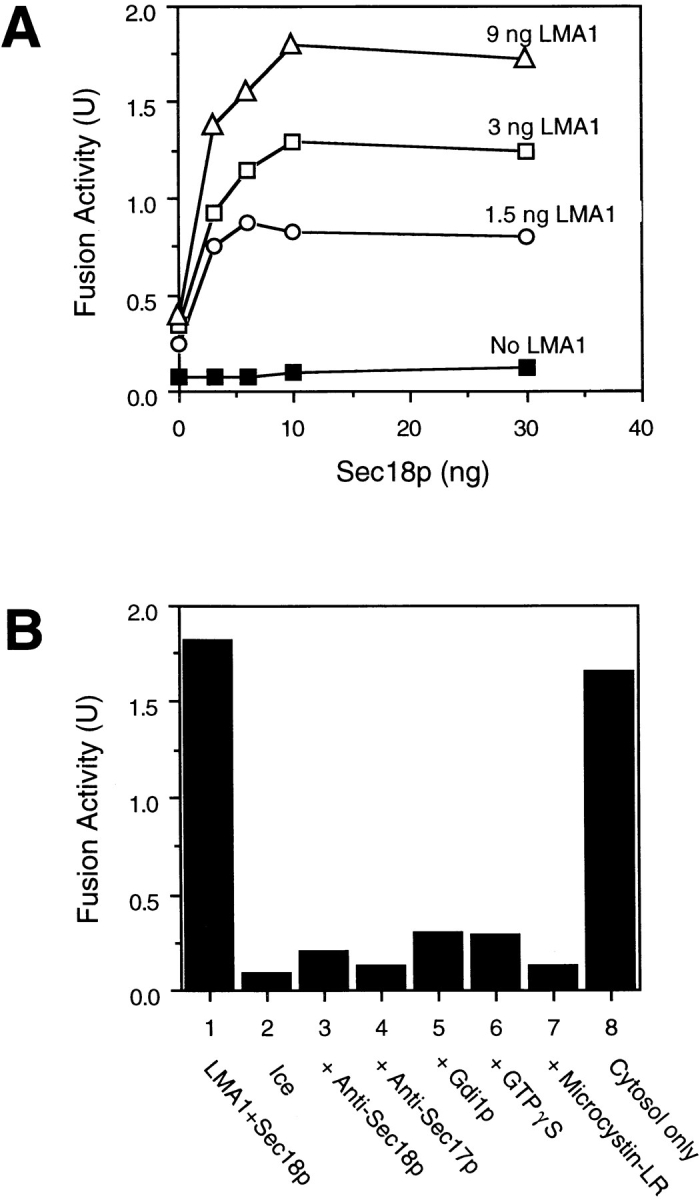
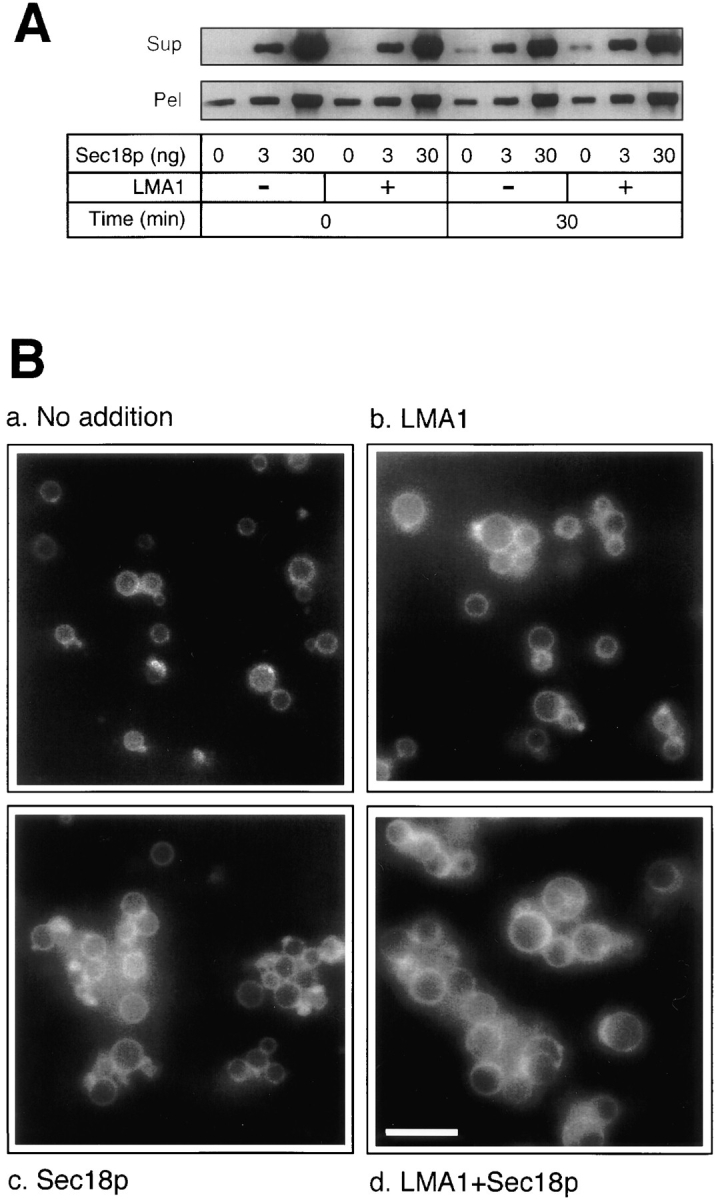
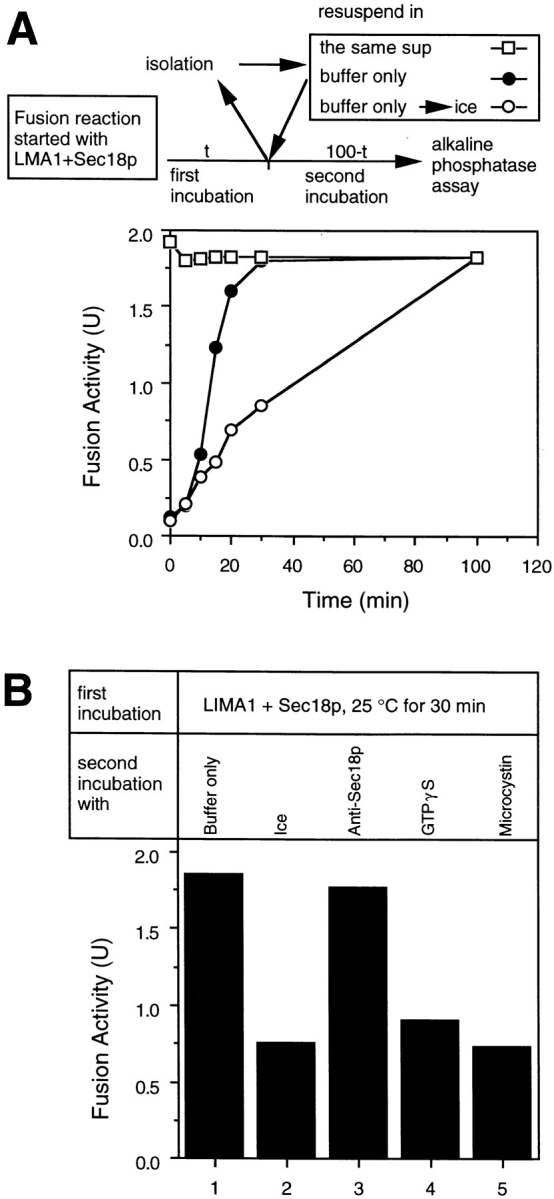
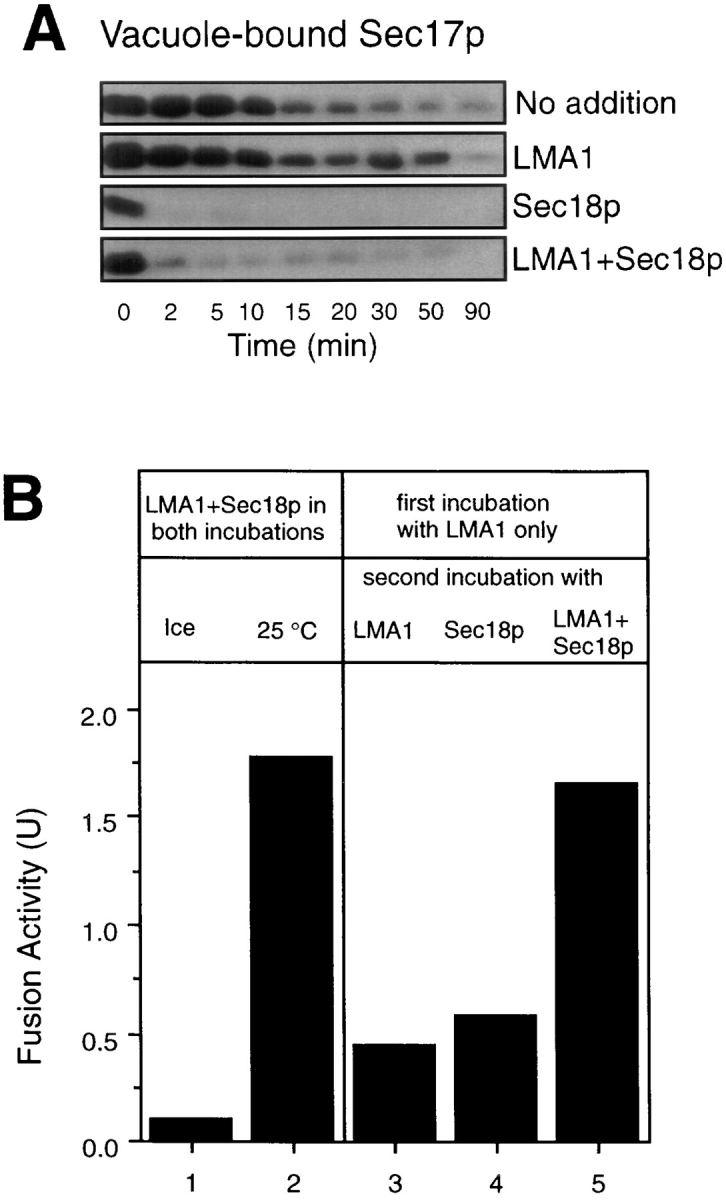
Early in S phase, the vacuole (lysosome) of Saccharomyces cerevisiae projects a stream of vesicles and membranous tubules into the bud where they fuse and establish the daughter vacuole. This inheritance reaction can be studied in vitro with isolated vacuoles. Rapid and efficient homotypic fusion between salt-washed vacuoles requires the addition of only two purified soluble proteins, Sec18p (NSF) and LMA1, a novel heterodimer with a thioredoxin subunit. We now report the identity of the second subunit of LMA1 as I(B)2, a previously identified cytosolic inhibitor of vacuolar proteinase B. Both subunits are needed for efficient vacuole inheritance in vivo and for the LMA1 activity in cell extracts. Each subunit acts via a novel mechanism, as the thioredoxin subunit is not acting through redox chemistry and LMA1 is still needed for the fusion of vacuoles which do not contain proteinase B. Both Sec18p and LMA1 act at an early stage of the in vitro reaction. Though LMA1 does not stimulate Sec18p-mediated Sec17p release, LMA1 cannot fulfill its function before Sec18p. Upon Sec17p/Sec18p action, vacuoles become labile but are rapidly stabilized by LMA1. The action of LMA1 and Sec18p is thus coupled and ordered. These data establish LMA1 as a novel factor in trafficking of yeast vacuoles.
在S期早期,酿酒酵母的液泡(溶酶体)会向芽中投射出一股囊泡和膜性小管,它们在芽中融合并形成子液泡。这种遗传反应可以用分离出的液泡在体外进行研究。盐洗后的液泡之间快速而高效的同型融合仅需添加两种纯化的可溶性蛋白,即Sec18p(NSF)和LMA1,LMA1是一种具有硫氧还蛋白亚基的新型异二聚体。我们现在报告LMA1的第二个亚基是I(B)2,它是先前鉴定出的液泡蛋白酶B的胞质抑制剂。这两个亚基对于体内有效的液泡遗传以及细胞提取物中的LMA1活性都是必需的。每个亚基都通过一种新机制起作用,因为硫氧还蛋白亚基并非通过氧化还原化学起作用,而且对于不含蛋白酶B的液泡融合,LMA1仍然是必需的。Sec18p和LMA1都在体外反应的早期阶段起作用。虽然LMA1不会刺激Sec18p介导的Sec17p释放,但在Sec18p发挥作用之前,LMA1无法履行其功能。在Sec17p/Sec18p作用后,液泡变得不稳定,但会迅速被LMA1稳定。因此,LMA1和Sec18p的作用是相互关联且有序的。这些数据确立了LMA1是酵母液泡运输中的一个新因子。