Cheung M, Akabas M H
Center for Molecular Recognition, College of Physicians and Surgeons, Columbia University, New York 10032, USA.
J Gen Physiol. 1997 Mar;109(3):289-99. doi: 10.1085/jgp.109.3.289.
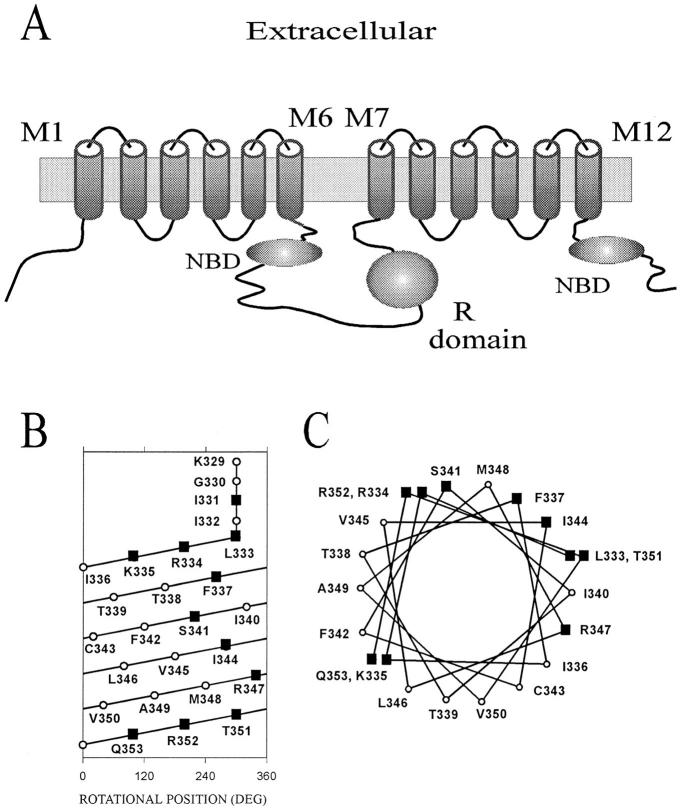
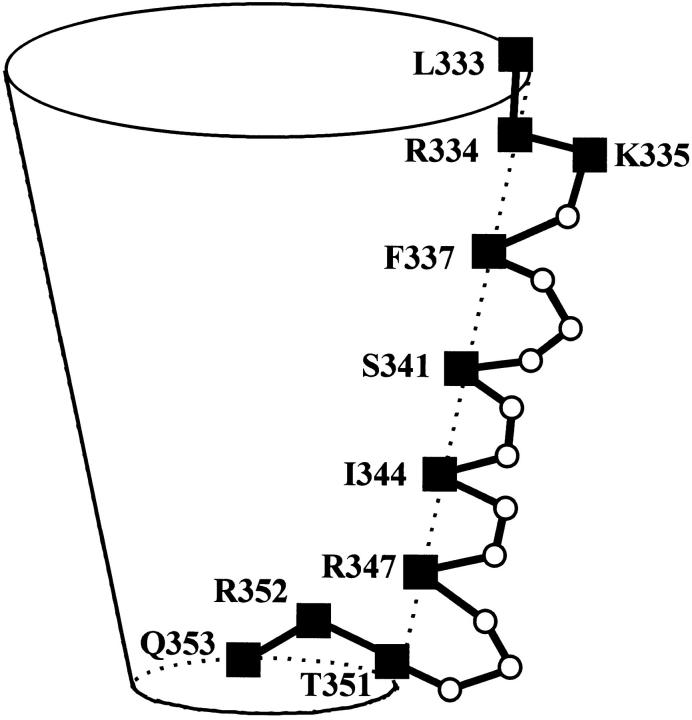
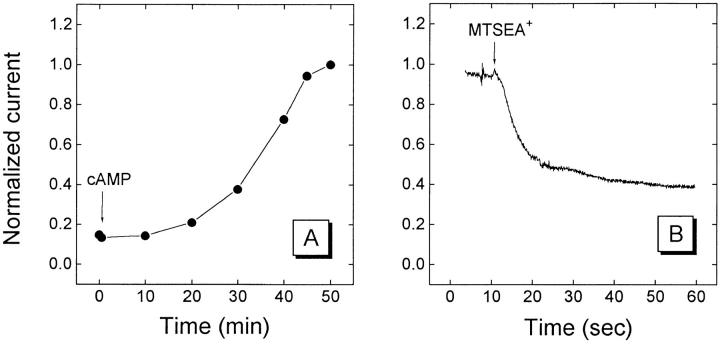
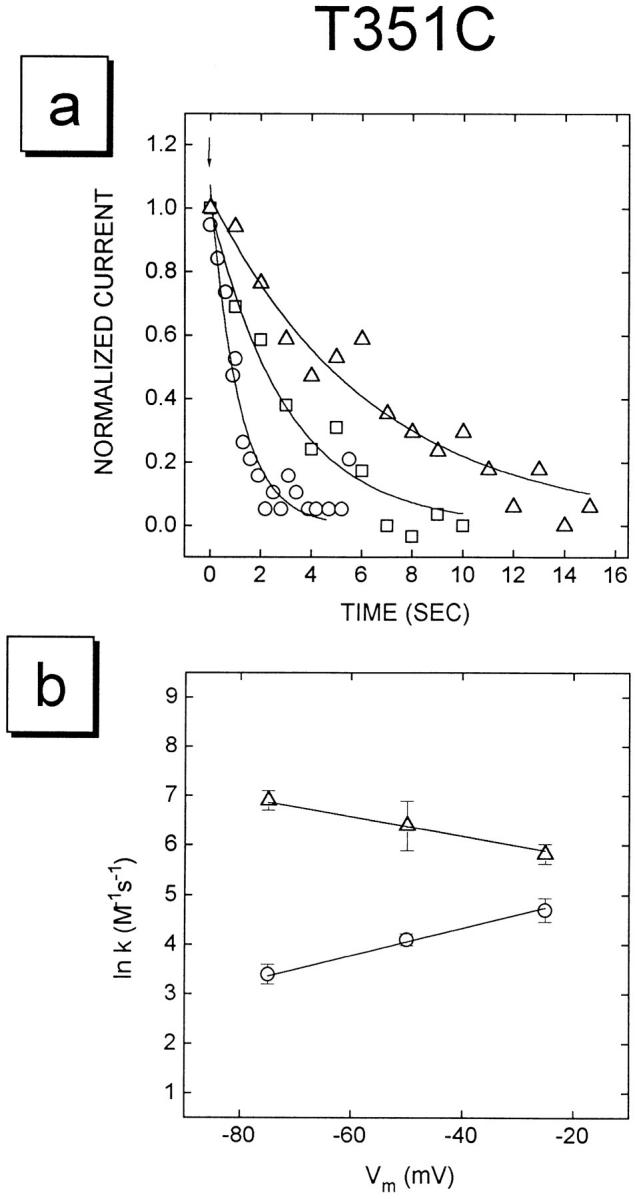
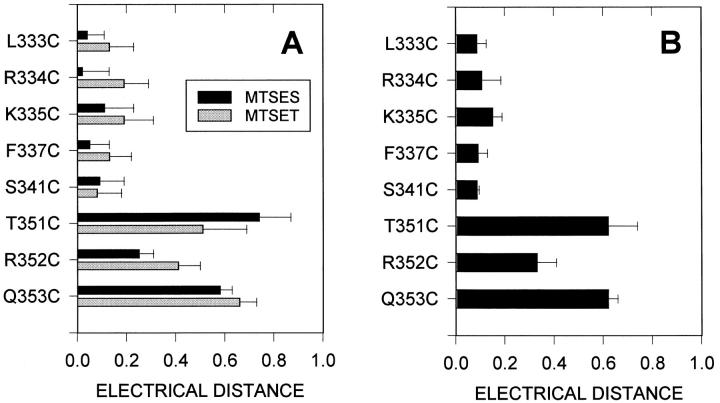
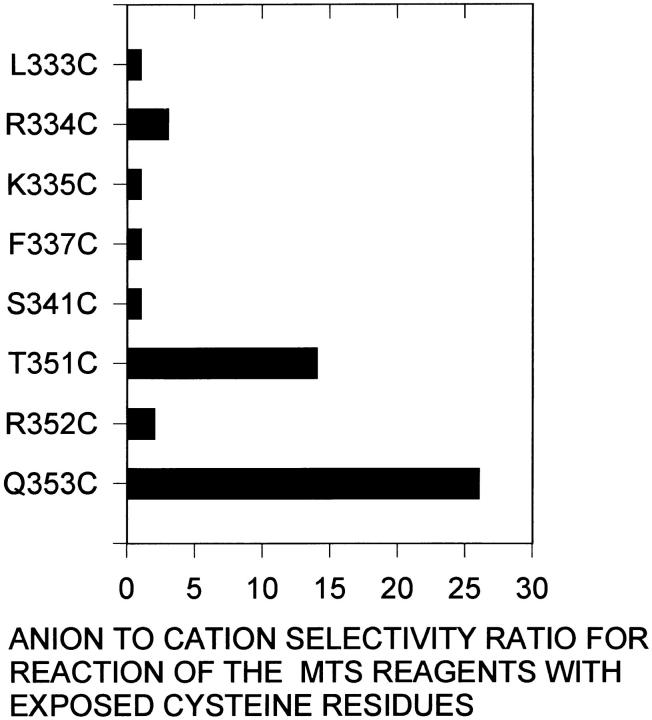
The cystic fibrosis transmembrane conductance regulator forms an anion-selective channel; the site and mechanism of charge selectivity is unknown. We previously reported that cysteines substituted, one at a time, for Ile331, Leu333, Arg334, Lys335, Phe337, Ser341, Ile344, Arg347, Thr351, Arg352, and Gln353, in and flanking the sixth membrane-spanning segment (M6), reacted with charged, sulfhydryl-specific, methanethiosulfonate (MTS) reagents. We inferred that these residues are on the water-accessible surface of the protein and may line the ion channel. We have now measured the voltage-dependence of the reaction rates of the MTS reagents with the accessible, engineering cysteines. By comparing the reaction rates of negatively and positively charged MTS reagents with these cysteines, we measured the extent of anion selectivity from the extracellular end of the channel to eight of the accessible residues. We show that the major site determining anion vs. cation selectivity is near the cytoplasmic end of the channel; it favors anions by approximately 25-fold and may involve the residues Arg347 and Arg 352. From the voltage dependence of the reaction rates, we calculated the electrical distance to the accessible residues. For the residues from Leu333 to Ser341 the electrical distance is not significantly different than zero; it is significantly different than zero for the residues Thr351 to Gln353. The maximum electrical distance measured was 0.6 suggesting that the channel extends more cytoplasmically and may include residues flanking the cytoplasmic end of the M6 segment. Furthermore, the electrical distance calculations indicate that R352C is closer to the extracellular end of the channel than either of the adjacent residues. We speculate that the cytoplasmic end of the M6 segment may loop back into the channel narrowing the lumen and thereby forming both the major resistance to current flow and the anion-selectivity filter.
囊性纤维化跨膜传导调节因子形成一个阴离子选择性通道;电荷选择性的位点和机制尚不清楚。我们之前报道过,在第六个跨膜片段(M6)及其侧翼,一次用一个半胱氨酸取代异亮氨酸331、亮氨酸333、精氨酸334、赖氨酸335、苯丙氨酸337、丝氨酸341、异亮氨酸344、精氨酸347、苏氨酸351、精氨酸352和谷氨酰胺353,这些半胱氨酸会与带电荷的、巯基特异性的甲硫基磺酸盐(MTS)试剂发生反应。我们推断这些残基位于蛋白质的水可及表面,可能构成离子通道的内壁。我们现在测量了MTS试剂与可及的工程化半胱氨酸反应速率的电压依赖性。通过比较带负电荷和正电荷的MTS试剂与这些半胱氨酸的反应速率,我们测量了从通道细胞外端到八个可及残基的阴离子选择性程度。我们表明,决定阴离子与阳离子选择性的主要位点靠近通道的细胞质端;它对阴离子的偏好约为25倍,可能涉及残基精氨酸347和精氨酸352。根据反应速率的电压依赖性,我们计算了到可及残基的电距离。对于从亮氨酸333到丝氨酸341的残基,电距离与零没有显著差异;对于从苏氨酸351到谷氨酰胺353的残基,电距离与零有显著差异。测量到的最大电距离为0.6,这表明通道在细胞质方向延伸得更多,可能包括M6片段细胞质端侧翼的残基。此外,电距离计算表明,R352C比任何一个相邻残基更靠近通道的细胞外端。我们推测M6片段的细胞质端可能回折到通道中,使管腔变窄,从而形成对电流流动的主要阻力和阴离子选择性过滤器。