Philips J, Herskowitz I
Department of Biochemistry and Biophysics, Programs in Genetics and Cell Biology, San Francisco, California 94143-0448, USA.
J Cell Biol. 1997 Sep 8;138(5):961-74. doi: 10.1083/jcb.138.5.961.
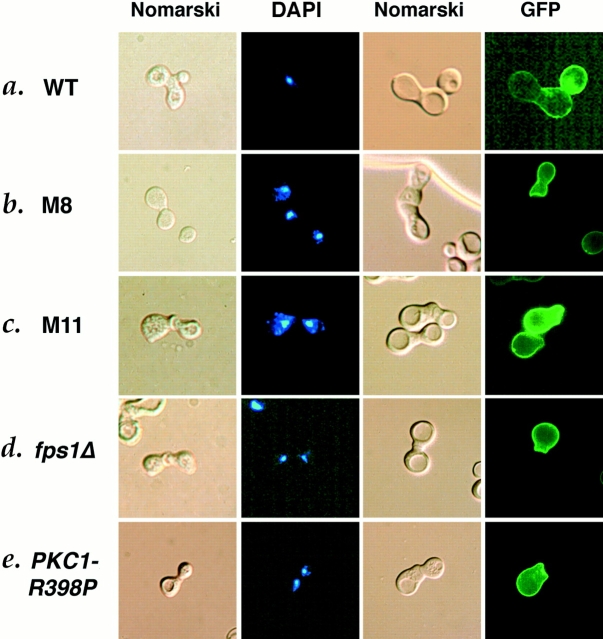
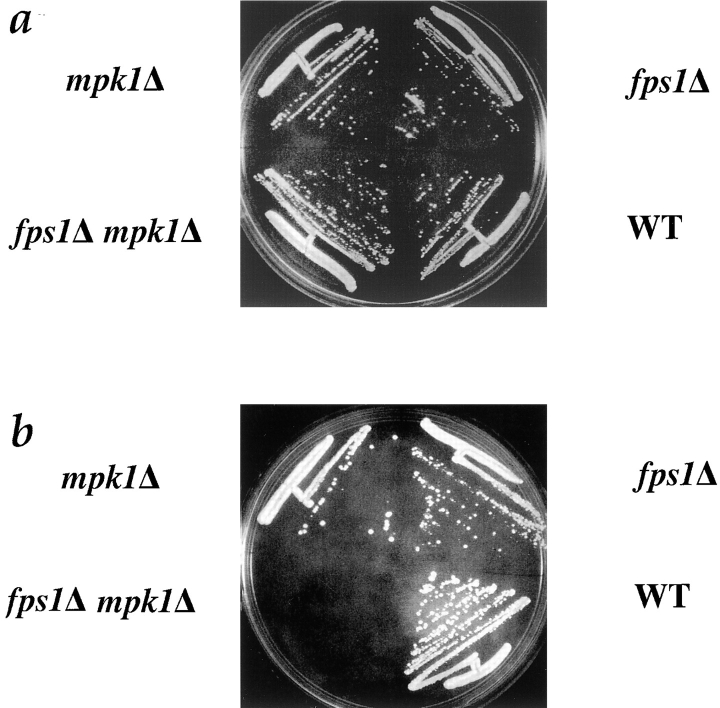
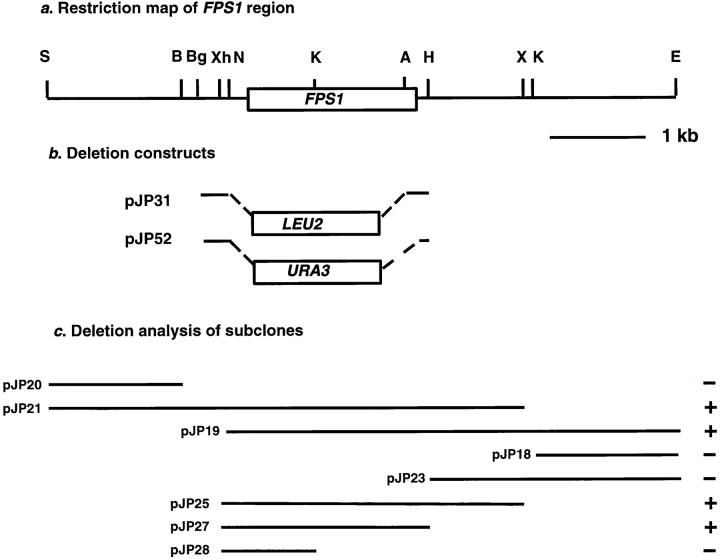
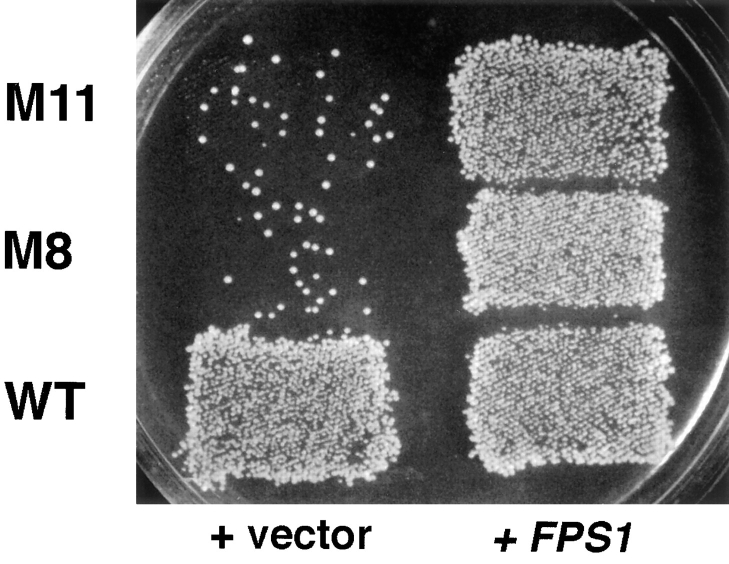
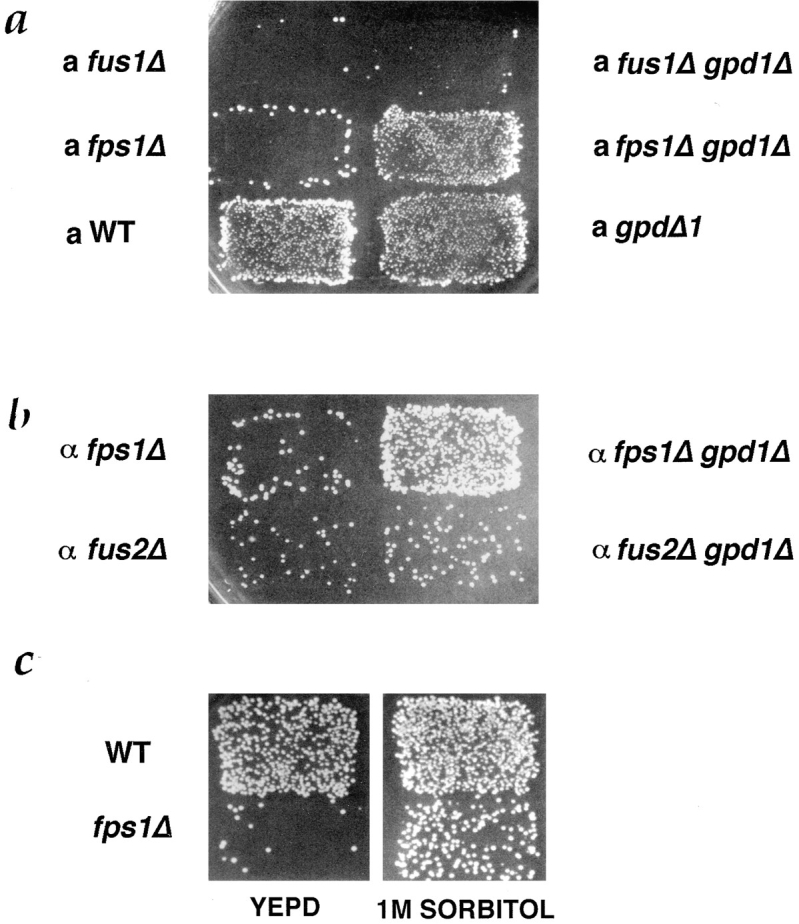
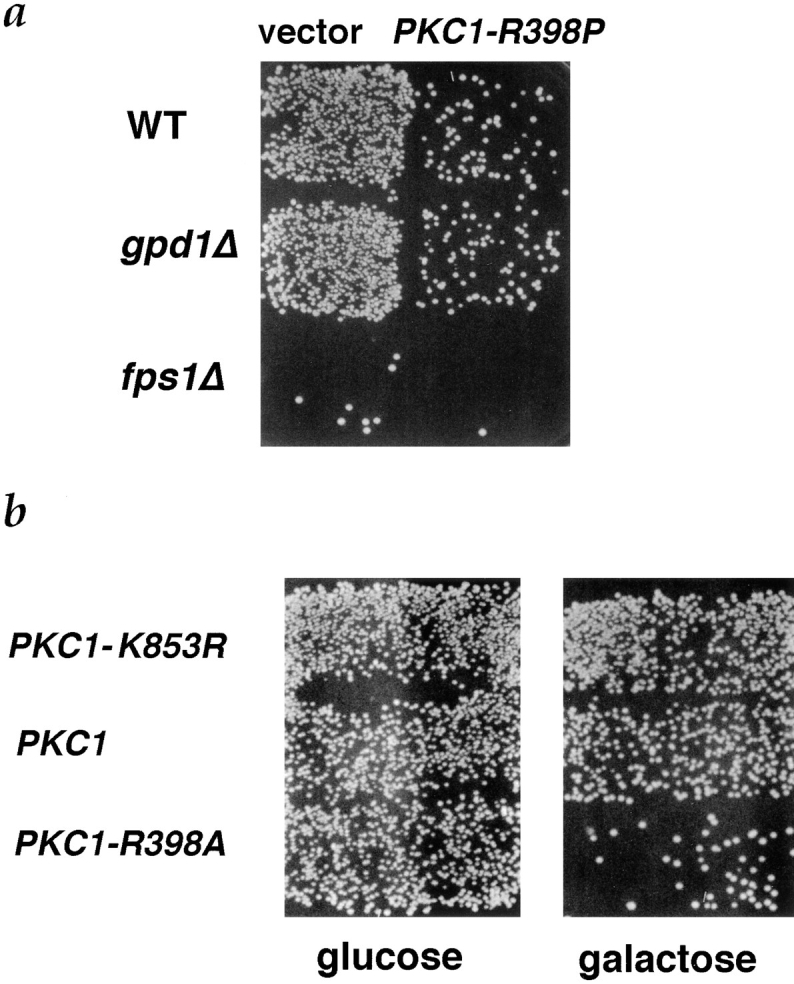
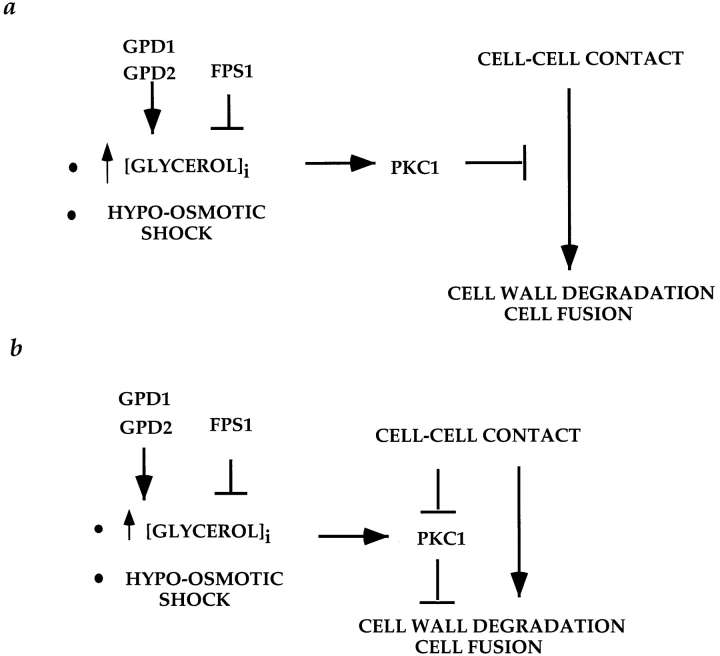
Successful zygote formation during yeast mating requires cell fusion of the two haploid mating partners. To ensure that cells do not lyse as they remodel their cell wall, the fusion event is both temporally and spatially regulated: the cell wall is degraded only after cell-cell contact and only in the region of cell-cell contact. To understand how cell fusion is regulated, we identified mutants defective in cell fusion based upon their defect in mating to a fus1 fus2 strain (Chenevert, J., N. Valtz, and I. Herskowitz. 1994. Genetics 136:1287-1297). Two of these cell fusion mutants are defective in the FPS1 gene, which codes for a glycerol facilitator (Luyten, K., J. Albertyn, W.F. Skibbe, B.A. Prior, J. Ramos, J.M. Thevelein, and S. Hohmann. 1995. EMBO [Eur. Mol. Biol. Organ.] J. 14:1360-1371). To determine whether inability to maintain osmotic balance accounts for the defect in cell fusion in these mutants, we analyzed the behavior of an fps1Delta mutant with reduced intracellular glycerol levels because of a defect in the glycerol-3-phosphate dehydrogenase (GPD1) gene (Albertyn, J., S. Hohmann, J.M. Thevelein, and B.A. Prior. 1994. Mol. Cell. Biol. 14:4135-4144): deletion of GPD1 partially suppressed the cell fusion defect of fps1 mutants. In contrast, overexpression of GPD1 exacerbated the defect. The fusion defect could also be partially suppressed by 1 M sorbitol. These observations indicate that the fusion defect of fps1 mutants results from inability to regulate osmotic balance and provide evidence that the osmotic state of the cell can regulate fusion. We have also observed that mutants expressing hyperactive protein kinase C exhibit a cell fusion defect similar to that of fps1 mutants. We propose that Pkc1p regulates cell fusion in response to osmotic disequilibrium. Unlike fps1 mutants, fus1 and fus2 mutants are not influenced by expression of GPD1 or by 1 M sorbitol. Their fusion defect is thus unlikely to result from altered osmotic balance.
酵母交配过程中成功形成合子需要两个单倍体交配型细胞进行细胞融合。为确保细胞在重塑细胞壁时不会裂解,融合事件在时间和空间上均受到调控:细胞壁仅在细胞间接触后且仅在细胞间接触区域被降解。为了解细胞融合是如何被调控的,我们基于与fus1 fus2菌株交配存在缺陷,鉴定出了细胞融合缺陷的突变体(Chenevert, J., N. Valtz, and I. Herskowitz. 1994. Genetics 136:1287 - 1297)。其中两个细胞融合突变体在FPS1基因上存在缺陷,该基因编码一种甘油转运蛋白(Luyten, K., J. Albertyn, W.F. Skibbe, B.A. Prior, J. Ramos, J.M. Thevelein, and S. Hohmann. 1995. EMBO [Eur. Mol. Biol. Organ.] J. 14:1360 - 1371)。为确定无法维持渗透平衡是否导致了这些突变体的细胞融合缺陷,我们分析了一个fps1Delta突变体的行为,该突变体由于甘油 - 3 - 磷酸脱氢酶(GPD1)基因缺陷而细胞内甘油水平降低(Albertyn, J., S. Hohmann, J.M. Thevelein, and B.A. Prior. 1994. Mol. Cell. Biol. 14:4135 - 4144):删除GPD1部分抑制了fps1突变体的细胞融合缺陷。相反,GPD1的过表达加剧了该缺陷。1 M山梨醇也可部分抑制融合缺陷。这些观察结果表明,fps1突变体的融合缺陷是由于无法调节渗透平衡导致的,并提供了细胞的渗透状态可调节融合的证据。我们还观察到,表达活性过高的蛋白激酶C的突变体表现出与fps1突变体类似的细胞融合缺陷。我们提出,Pkc1p响应渗透失衡来调节细胞融合。与fps1突变体不同,fus1和fus2突变体不受GPD1表达或1 M山梨醇的影响。因此,它们的融合缺陷不太可能是由渗透平衡改变导致的。