Mauerer U R, Boulpaep E L, Segal A S
Department of Cellular and Molecular Physiology, Yale University School of Medicine, New Haven, Connecticut 06520, USA.
J Gen Physiol. 1998 Jan;111(1):139-60. doi: 10.1085/jgp.111.1.139.
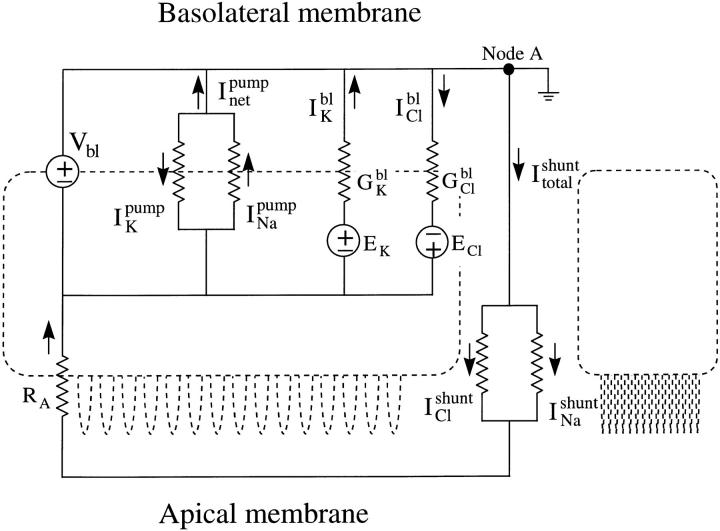
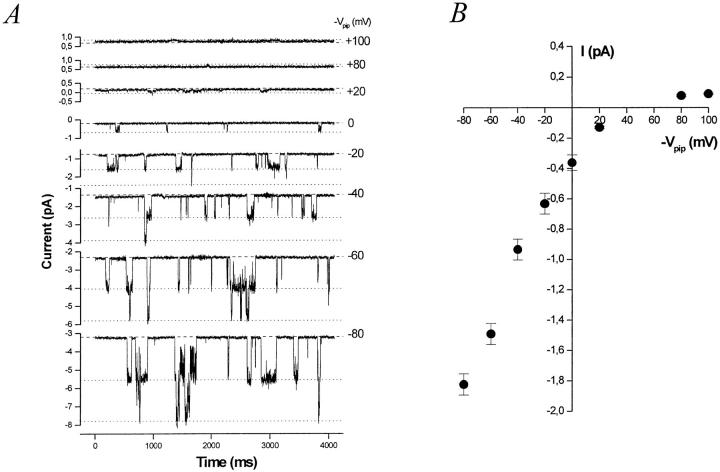
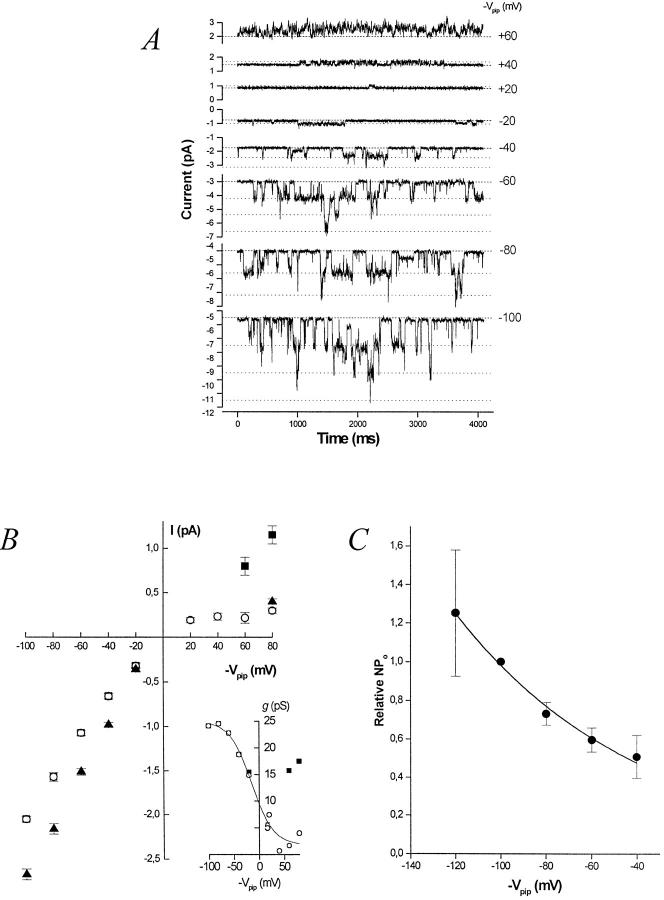
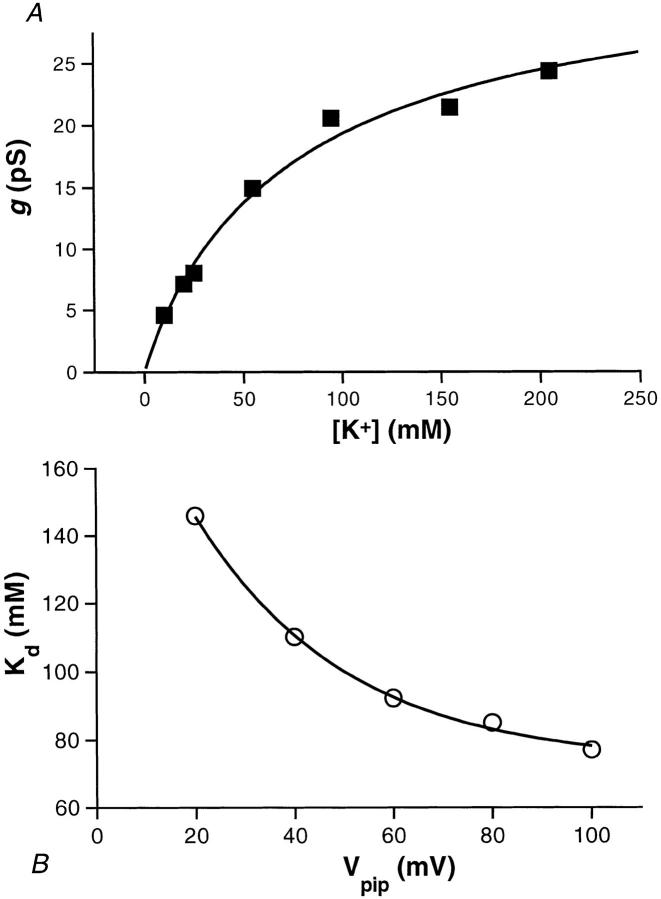
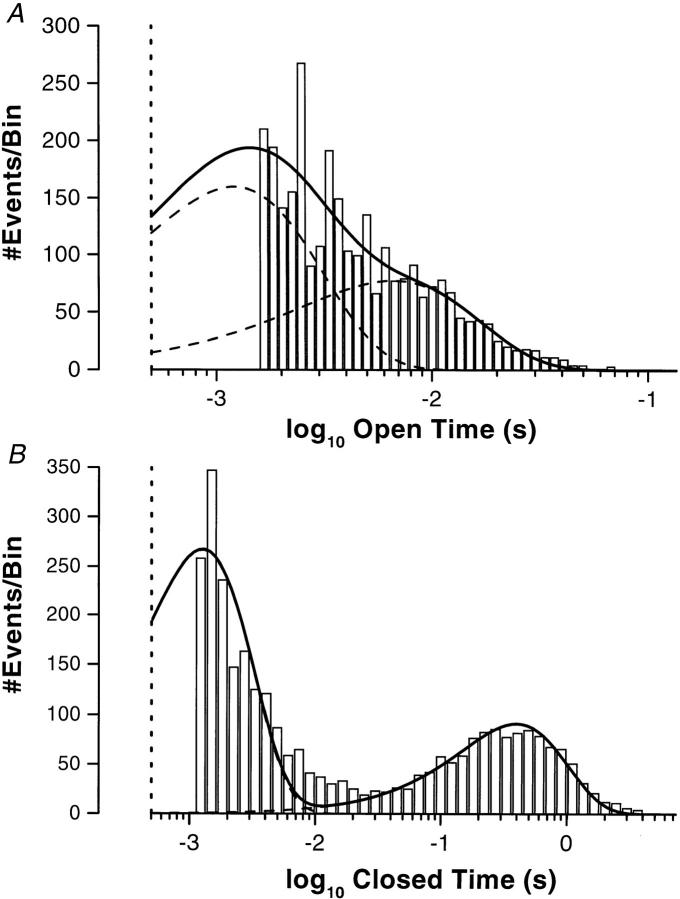
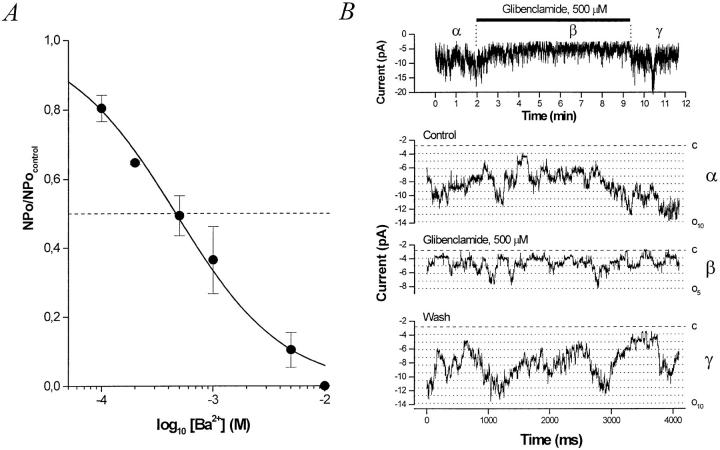
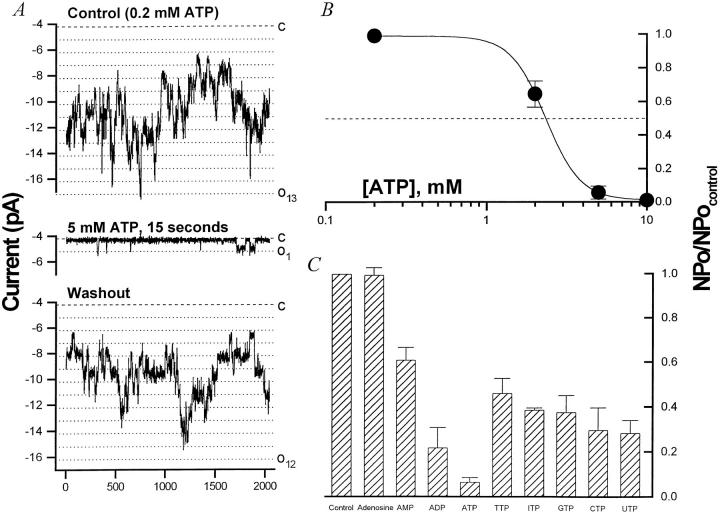
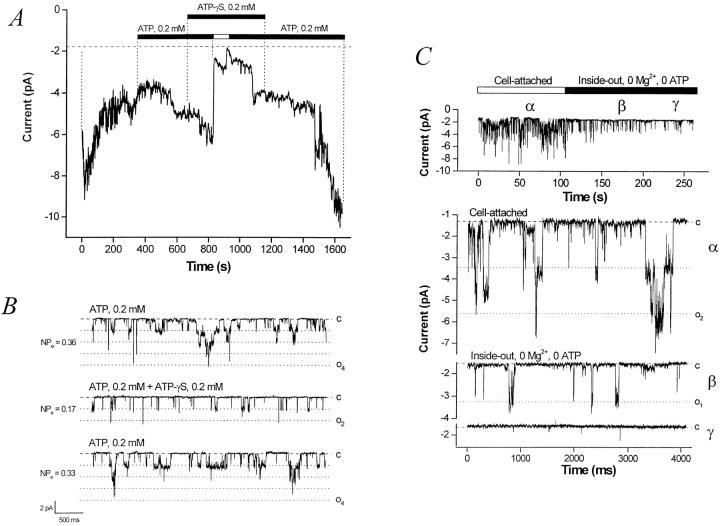
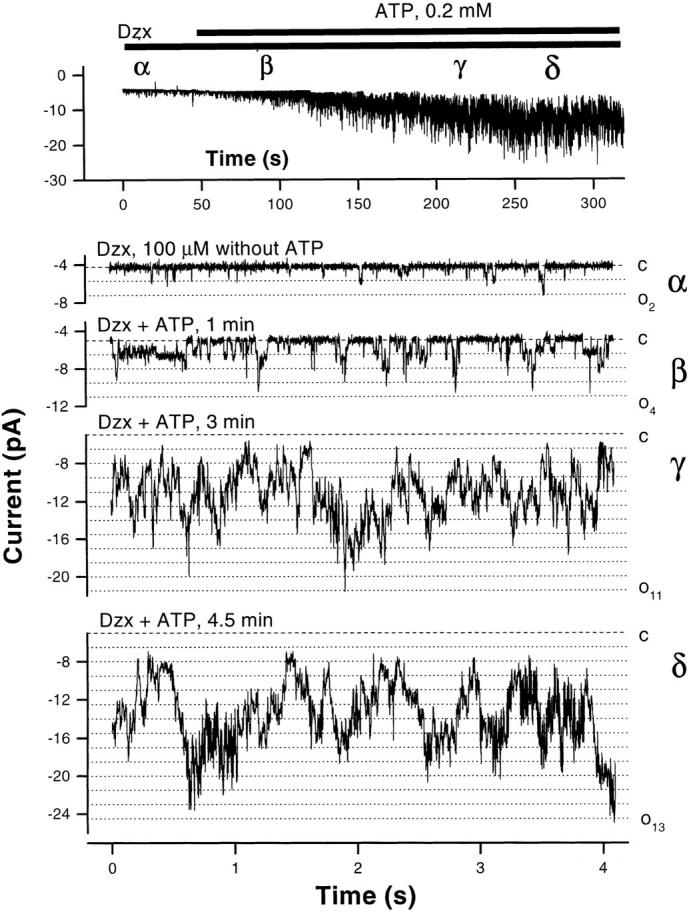
The potassium conductance of the basolateral membrane (BLM) of proximal tubule cells is a critical regulator of transport since it is the major determinant of the negative cell membrane potential and is necessary for pump-leak coupling to the Na+,K+-ATPase pump. Despite this pivotal physiological role, the properties of this conductance have been incompletely characterized, in part due to difficulty gaining access to the BLM. We have investigated the properties of this BLM K+ conductance in dissociated, polarized Ambystoma proximal tubule cells. Nearly all seals made on Ambystoma cells contained inward rectifier K+ channels (gammaslope, in = 24.5 +/- 0.6 pS, gammachord, out = 3.7 +/- 0.4 pS). The rectification is mediated in part by internal Mg2+. The open probability of the channel increases modestly with hyperpolarization. The inward conducting properties are described by a saturating binding-unbinding model. The channel conducts Tl+ and K+, but there is no significant conductance for Na+, Rb+, Cs+, Li+, NH4+, or Cl-. The channel is inhibited by barium and the sulfonylurea agent glibenclamide, but not by tetraethylammonium. Channel rundown typically occurs in the absence of ATP, but cytosolic addition of 0. 2 mM ATP (or any hydrolyzable nucleoside triphosphate) sustains channel activity indefinitely. Phosphorylation processes alone fail to sustain channel activity. Higher doses of ATP (or other nucleoside triphosphates) reversibly inhibit the channel. The K+ channel opener diazoxide opens the channel in the presence of 0.2 mM ATP, but does not alleviate the inhibition of millimolar doses of ATP. We conclude that this K+ channel is the major ATP-sensitive basolateral K+ conductance in the proximal tubule.
近端小管细胞基底外侧膜(BLM)的钾电导是转运的关键调节因子,因为它是细胞膜负电位的主要决定因素,也是泵 - 漏与Na +,K + -ATP酶泵耦合所必需的。尽管具有这一关键的生理作用,但这种电导的特性尚未完全明确,部分原因是难以接触到BLM。我们研究了分离的、极化的美西螈近端小管细胞中这种BLM钾电导的特性。几乎所有在美西螈细胞上形成的封接都包含内向整流钾通道(γ斜率,内向= 24.5±0.6 pS,γ弦,外向= 3.7±0.4 pS)。整流部分由细胞内Mg2 +介导。通道的开放概率随超极化适度增加。内向传导特性由饱和结合 - 解离模型描述。该通道传导Tl +和K +,但对Na +、Rb +、Cs +、Li +、NH4 +或Cl -没有明显的电导。该通道被钡和磺酰脲类药物格列本脲抑制,但不被四乙铵抑制。通道通常在没有ATP的情况下发生衰减,但胞质中添加0.2 mM ATP(或任何可水解的核苷三磷酸)可无限期维持通道活性。仅磷酸化过程不能维持通道活性。更高剂量的ATP(或其他核苷三磷酸)可逆地抑制该通道。钾通道开放剂二氮嗪在存在0.2 mM ATP的情况下打开通道,但不能减轻毫摩尔剂量ATP的抑制作用。我们得出结论,该钾通道是近端小管中主要的ATP敏感基底外侧钾电导。