Lenormand P, Brondello J M, Brunet A, Pouysségur J
Centre de Biochimie-Centre National de la Recherche Scientifique (CNRS) UMR 6543, Université de Nice, 06108 Nice, France.
J Cell Biol. 1998 Aug 10;142(3):625-33. doi: 10.1083/jcb.142.3.625.
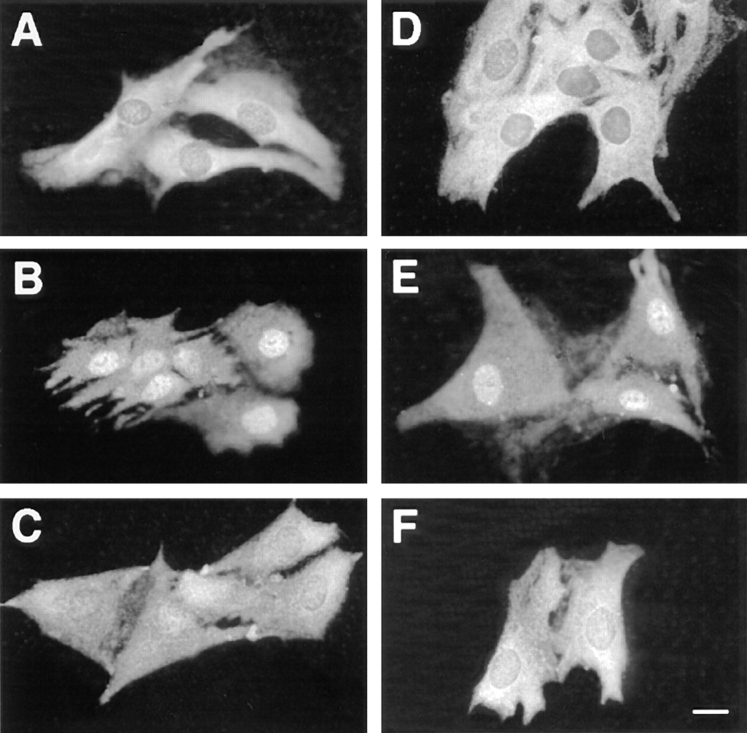
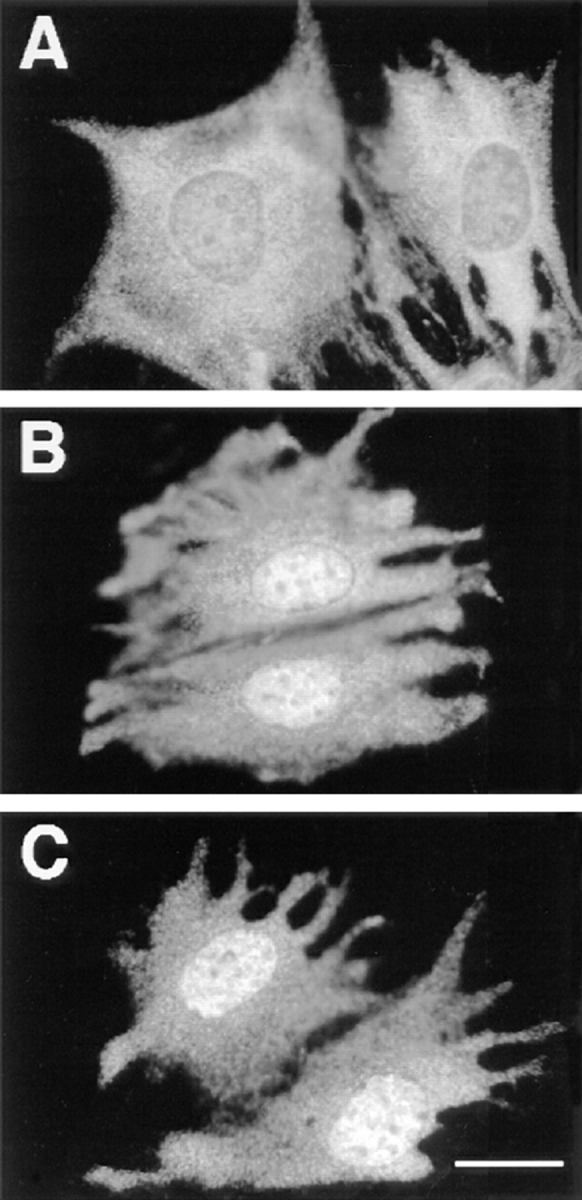
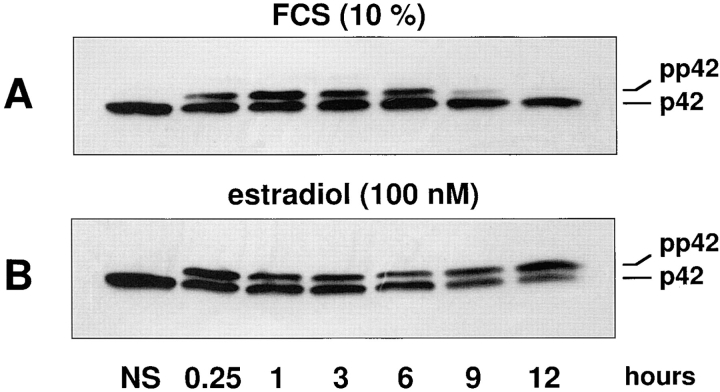
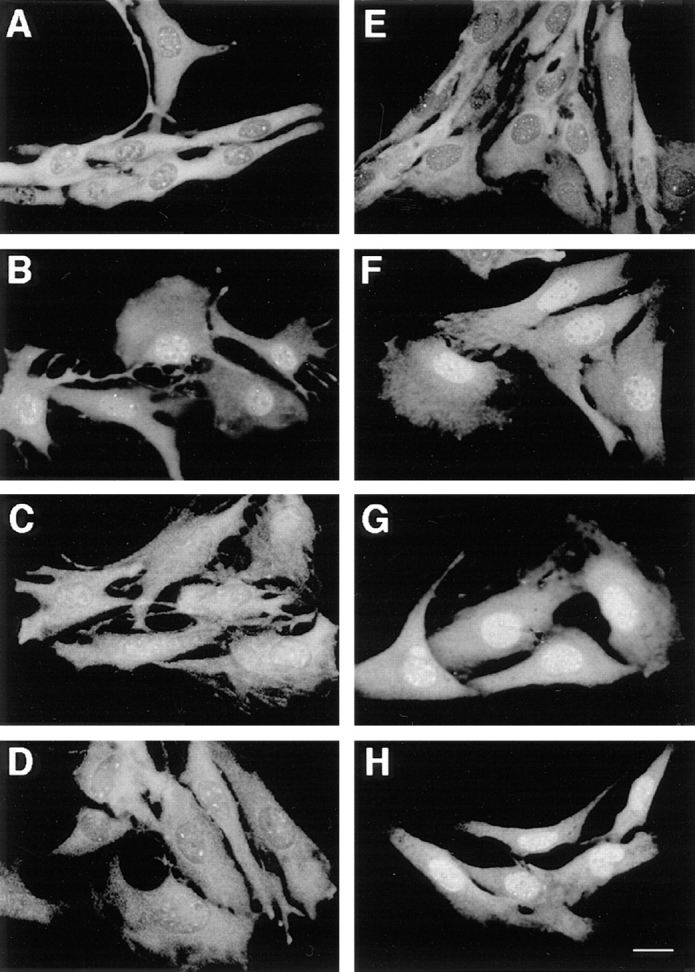
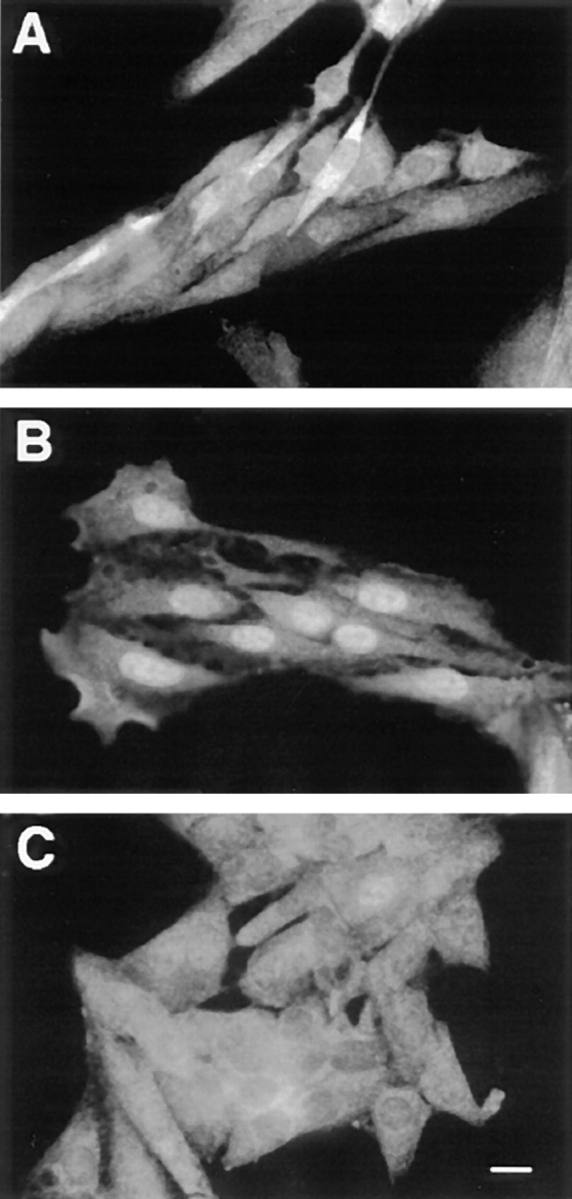
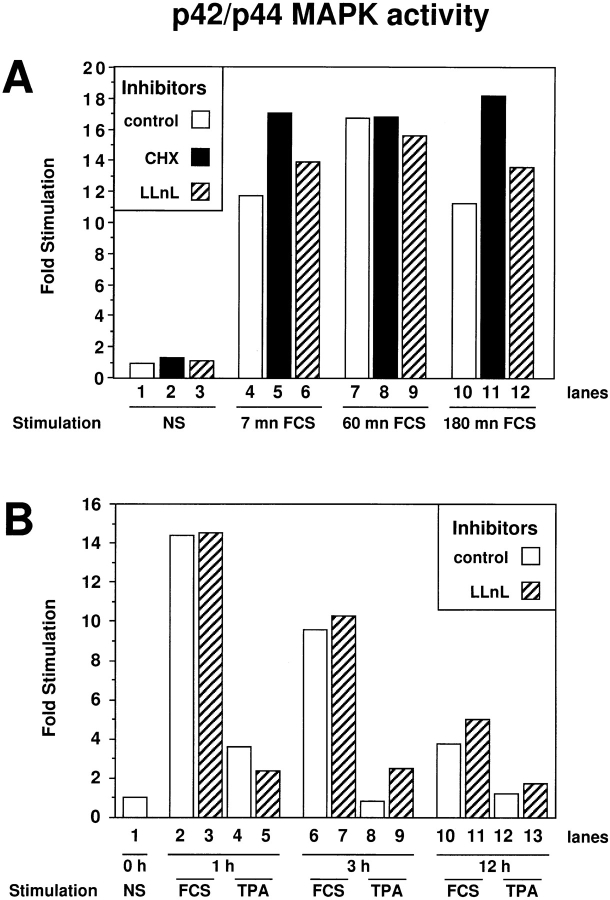
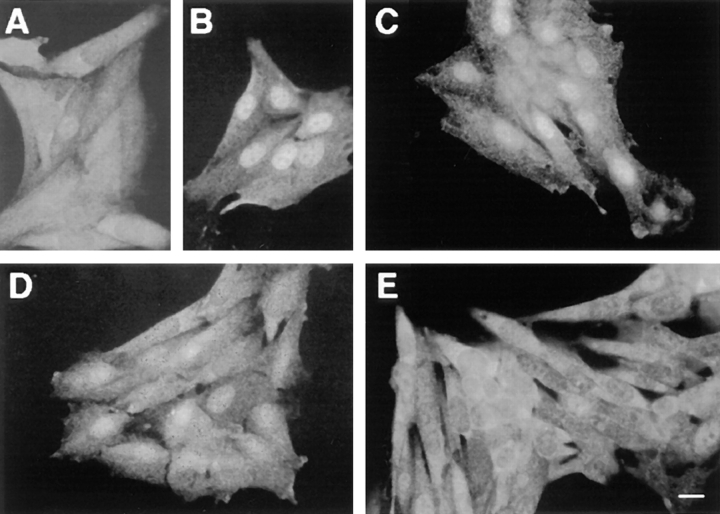
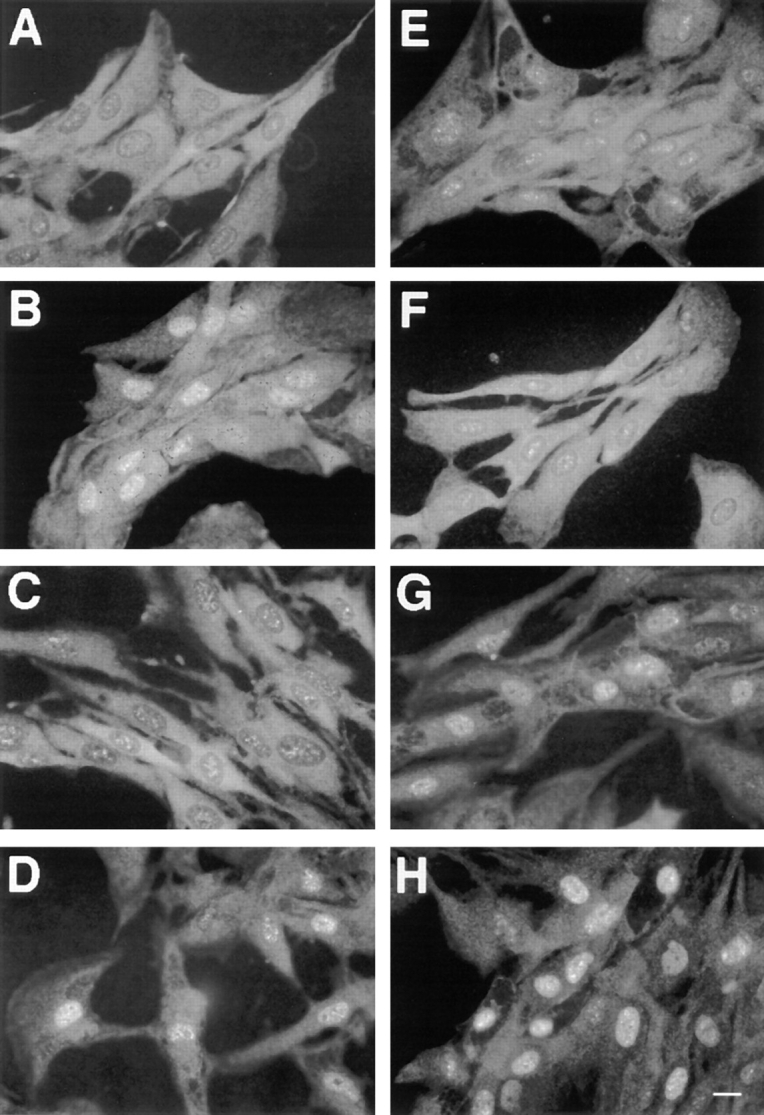
Mitogen-activated protein kinases (p42/p44 MAPK, also called Erk2 and Erk1) are key mediators of signal transduction from the cell surface to the nucleus. We have previously shown that the activation of p42/p44 MAPK required for transduction of mitogenic signaling is associated with a rapid nuclear translocation of these kinases. However, the means by which p42 and p44 MAPK translocate into the nucleus after cytoplasmic activation is still not understood and cannot simply be deduced from their protein sequences. In this study, we have demonstrated that activation of the p42/ p44 MAPK pathway was necessary and sufficient for triggering nuclear translocation of p42 and p44 MAPK. First, addition of the MEK inhibitor PD 98059, which blocks activation of the p42/p44 MAPK pathway, impedes the nuclear accumulation, whereas direct activation of the p42/p44 MAPK pathway by the chimera DeltaRaf-1:ER is sufficient to promote nuclear accumulation of p42/p44 MAPK. In addition, we have shown that this nuclear accumulation of p42/p44 MAPK required the neosynthesis of short-lived proteins. Indeed, inhibitors of protein synthesis abrogate nuclear accumulation in response to serum and accelerate p42/p44 MAPK nuclear efflux under conditions of persistent p42/p44 MAPK activation. In contrast, inhibition of targeted proteolysis by the proteasome synergistically potentiated p42/p44 MAPK nuclear localization by nonmitogenic agonists and markedly prolonged nuclear localization of p42/p44 MAPK after mitogenic stimulation. We therefore conclude that the MAPK nuclear translocation requires both activation of the p42/p44 MAPK module and neosynthesis of short-lived proteins that we postulate to be nuclear anchors.
丝裂原活化蛋白激酶(p42/p44 MAPK,也称为Erk2和Erk1)是从细胞表面到细胞核的信号转导的关键介质。我们之前已经表明,有丝分裂原信号转导所需的p42/p44 MAPK的激活与这些激酶的快速核转位有关。然而,p42和p44 MAPK在细胞质激活后转位到细胞核的方式仍不清楚,也不能简单地从它们的蛋白质序列中推断出来。在这项研究中,我们已经证明p42/p44 MAPK途径的激活对于触发p42和p44 MAPK的核转位是必要且充分的。首先,添加阻断p42/p44 MAPK途径激活的MEK抑制剂PD 98059会阻碍核积累,而嵌合体DeltaRaf-1:ER对p42/p44 MAPK途径的直接激活足以促进p42/p44 MAPK的核积累。此外,我们已经表明p42/p44 MAPK的这种核积累需要短寿命蛋白质的新合成。事实上,蛋白质合成抑制剂消除了对血清的核积累反应,并在持续的p42/p44 MAPK激活条件下加速了p42/p44 MAPK的核外流。相反,蛋白酶体对靶向蛋白水解的抑制通过非有丝分裂激动剂协同增强了p42/p44 MAPK的核定位,并在有丝分裂刺激后显著延长了p42/p44 MAPK的核定位。因此,我们得出结论,MAPK核转位既需要p42/p44 MAPK模块的激活,也需要我们假定为核锚的短寿命蛋白质的新合成。