Svitkina T M, Borisy G G
Laboratory of Molecular Biology, University of Wisconsin, Madison, Wisconsin 53706, USA.
J Cell Biol. 1999 May 31;145(5):1009-26. doi: 10.1083/jcb.145.5.1009.
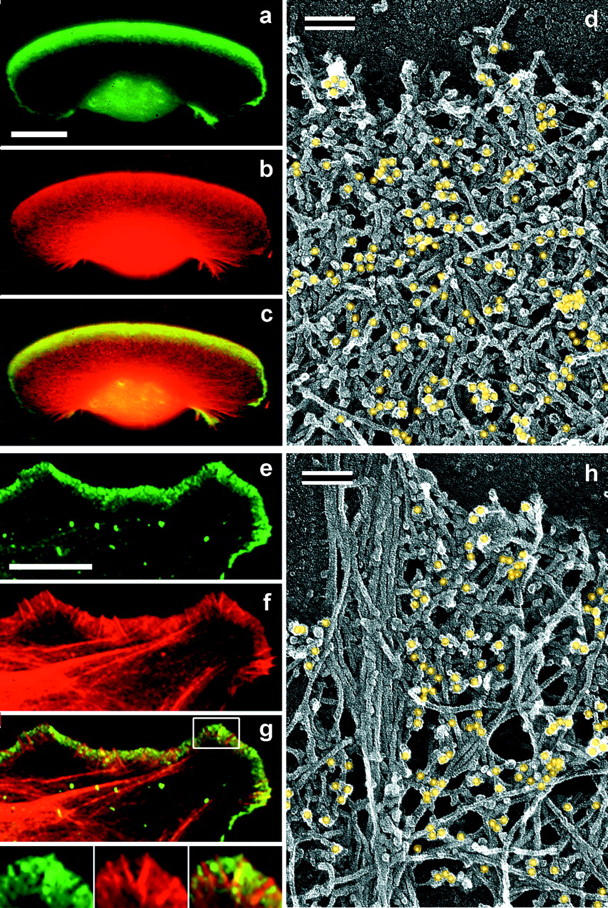
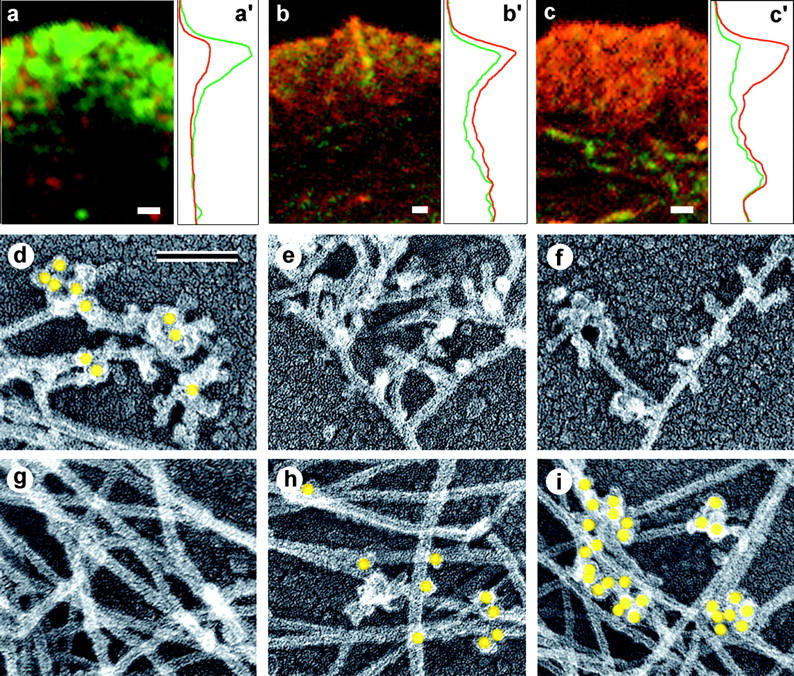
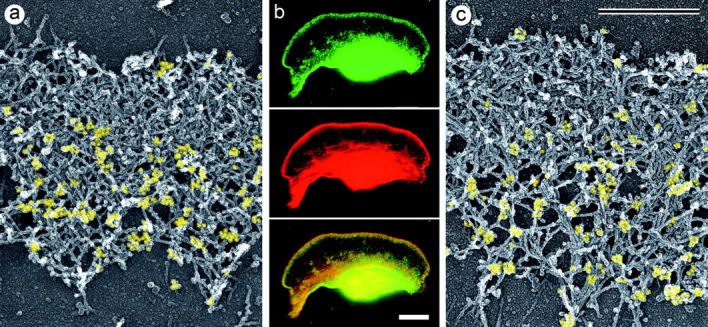
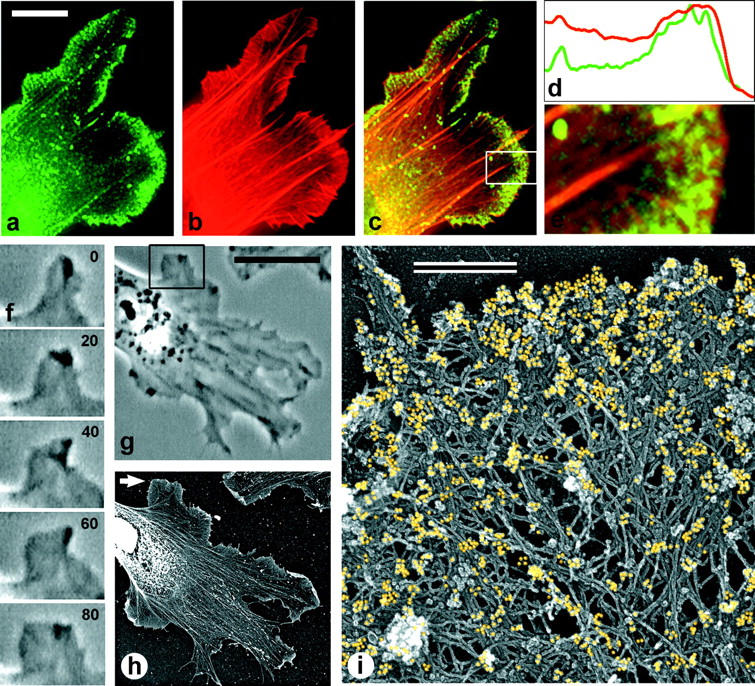
The leading edge (approximately 1 microgram) of lamellipodia in Xenopus laevis keratocytes and fibroblasts was shown to have an extensively branched organization of actin filaments, which we term the dendritic brush. Pointed ends of individual filaments were located at Y-junctions, where the Arp2/3 complex was also localized, suggesting a role of the Arp2/3 complex in branch formation. Differential depolymerization experiments suggested that the Arp2/3 complex also provided protection of pointed ends from depolymerization. Actin depolymerizing factor (ADF)/cofilin was excluded from the distal 0.4 micrometer++ of the lamellipodial network of keratocytes and in fibroblasts it was located within the depolymerization-resistant zone. These results suggest that ADF/cofilin, per se, is not sufficient for actin brush depolymerization and a regulatory step is required. Our evidence supports a dendritic nucleation model (Mullins, R.D., J.A. Heuser, and T.D. Pollard. 1998. Proc. Natl. Acad. Sci. USA. 95:6181-6186) for lamellipodial protrusion, which involves treadmilling of a branched actin array instead of treadmilling of individual filaments. In this model, Arp2/3 complex and ADF/cofilin have antagonistic activities. Arp2/3 complex is responsible for integration of nascent actin filaments into the actin network at the cell front and stabilizing pointed ends from depolymerization, while ADF/cofilin promotes filament disassembly at the rear of the brush, presumably by pointed end depolymerization after dissociation of the Arp2/3 complex.
非洲爪蟾角膜细胞和成纤维细胞片状伪足的前沿(约1微克)显示出肌动蛋白丝的广泛分支结构,我们将其称为树突状刷。单根细丝的尖端位于Y形连接处,Arp2/3复合体也定位于此处,这表明Arp2/3复合体在分支形成中发挥作用。差异解聚实验表明,Arp2/3复合体还能保护细丝尖端不发生解聚。肌动蛋白解聚因子(ADF)/丝切蛋白被排除在角膜细胞片状伪足网络的远端0.4微米++之外,在成纤维细胞中,它位于抗解聚区域内。这些结果表明,ADF/丝切蛋白本身不足以使肌动蛋白刷解聚,还需要一个调节步骤。我们的证据支持了一种关于片状伪足突出的树突状成核模型(Mullins, R.D., J.A. Heuser, and T.D. Pollard. 1998. Proc. Natl. Acad. Sci. USA. 95:6181-6186),该模型涉及分支肌动蛋白阵列的踏车行为,而不是单根细丝的踏车行为。在这个模型中,Arp2/3复合体和ADF/丝切蛋白具有拮抗活性。Arp2/3复合体负责将新生的肌动蛋白丝整合到细胞前端的肌动蛋白网络中,并稳定细丝尖端使其不发生解聚,而ADF/丝切蛋白则促进刷状结构后端的细丝解聚,推测是在Arp2/3复合体解离后通过尖端解聚实现的。