Altomare C, Bucchi A, Camatini E, Baruscotti M, Viscomi C, Moroni A, DiFrancesco D
Dipartimento di Fisiologia e Biochimica Generali, via Celoria 26, and INFM-Unità Milano Università, via Celoria 16, 20133 Milano, Italy.
J Gen Physiol. 2001 Jun;117(6):519-32. doi: 10.1085/jgp.117.6.519.
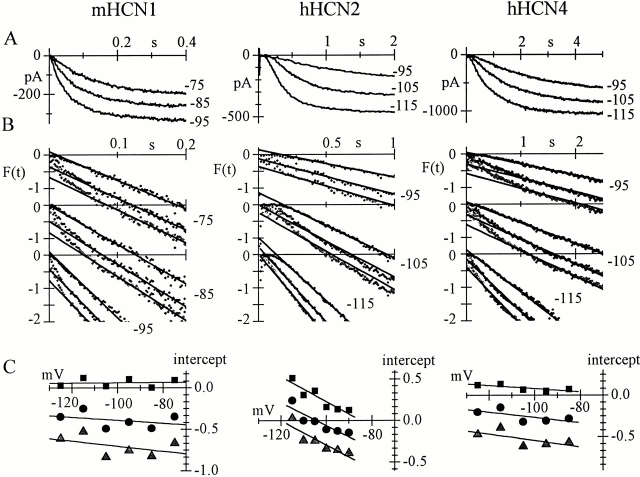
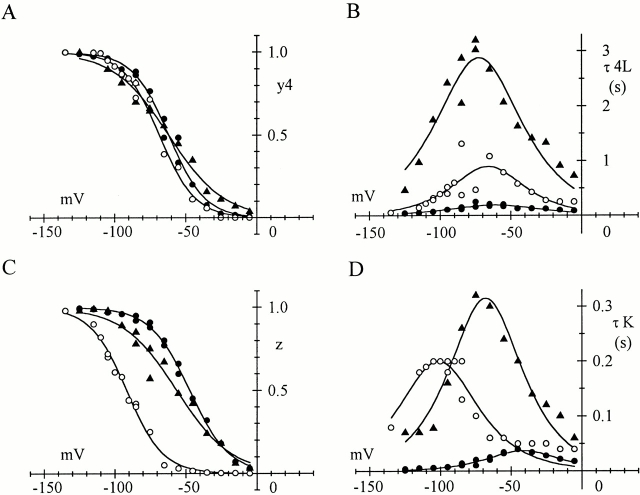
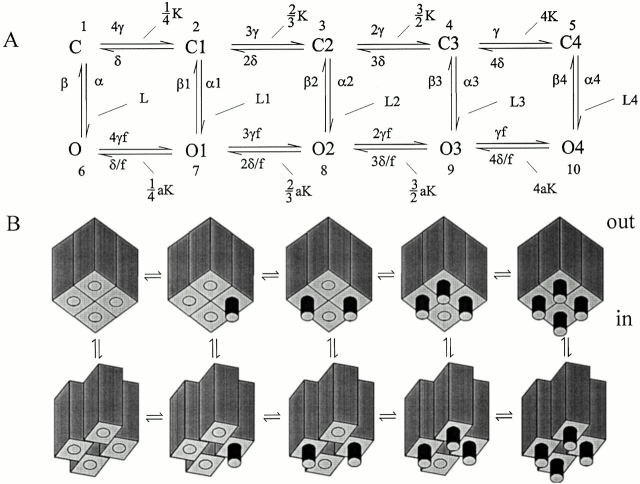
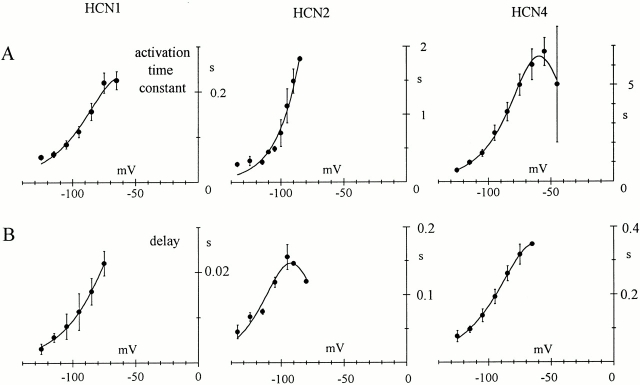
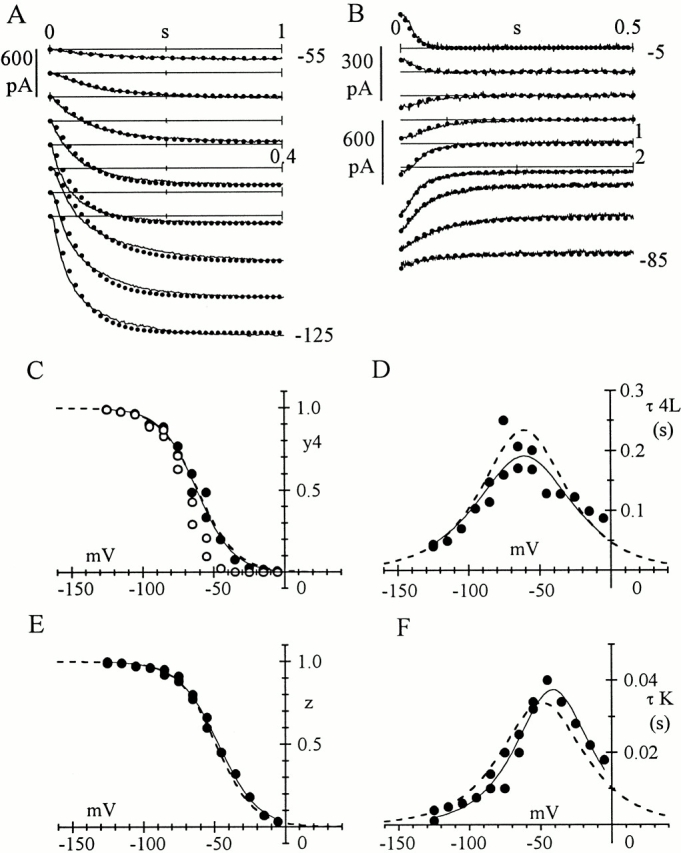
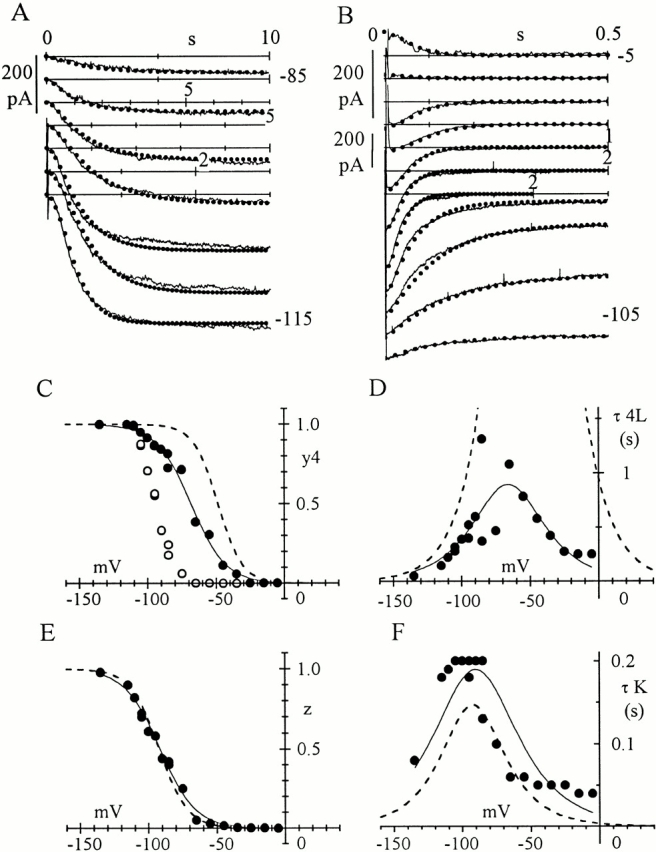
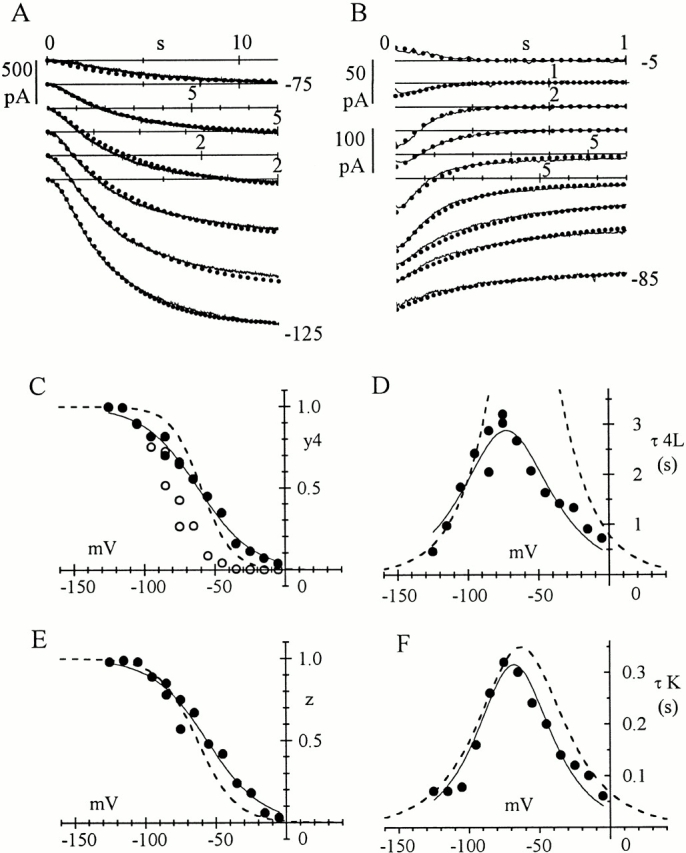
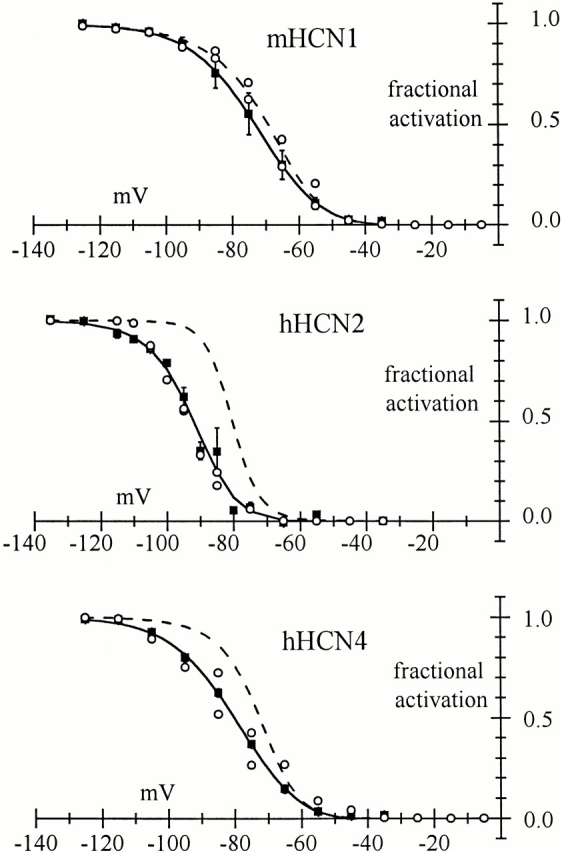
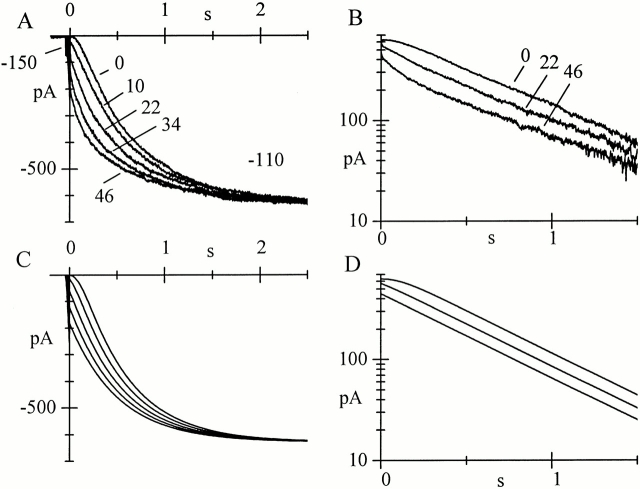
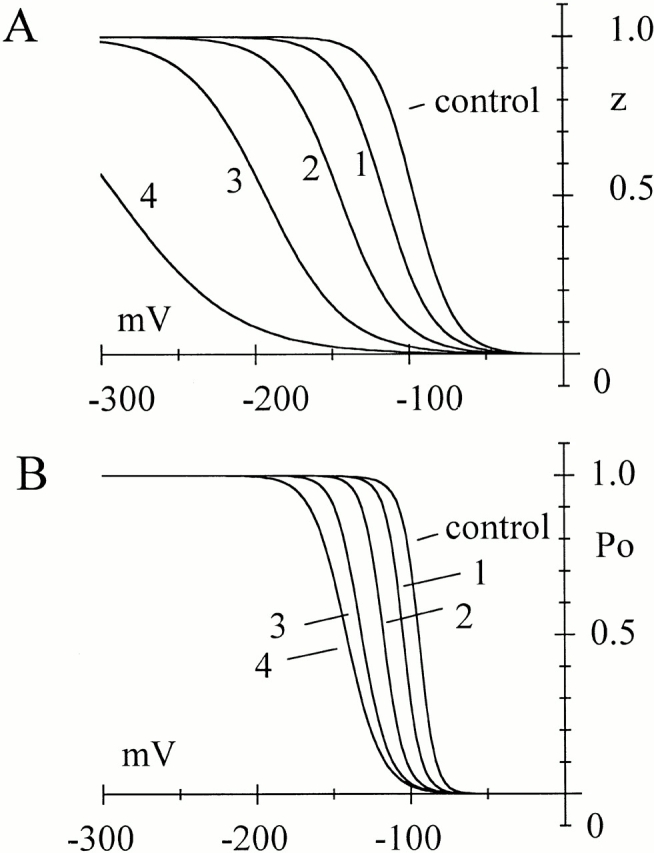
Hyperpolarization-activated (pacemaker) channels are dually gated by negative voltage and intracellular cAMP. Kinetics of native cardiac f-channels are not compatible with HH gating, and require closed/open multistate models. We verified that members of the HCN channel family (mHCN1, hHCN2, hHCN4) also have properties not complying with HH gating, such as sigmoidal activation and deactivation, activation deviating from fixed power of an exponential, removal of activation "delay" by preconditioning hyperpolarization. Previous work on native channels has indicated that the shifting action of cAMP on the open probability (Po) curve can be accounted for by an allosteric model, whereby cAMP binds more favorably to open than closed channels. We therefore asked whether not only cAMP-dependent, but also voltage-dependent gating of hyperpolarization-activated channels could be explained by an allosteric model. We hypothesized that HCN channels are tetramers and that each subunit comprises a voltage sensor moving between "reluctant" and "willing" states, whereas voltage sensors are independently gated by voltage, channel closed/open transitions occur allosterically. These hypotheses led to a multistate scheme comprising five open and five closed channel states. We estimated model rate constants by fitting first activation delay curves and single exponential time constant curves, and then individual activation/deactivation traces. By simply using different sets of rate constants, the model accounts for qualitative and quantitative aspects of voltage gating of all three HCN isoforms investigated, and allows an interpretation of the different kinetic properties of different isoforms. For example, faster kinetics of HCN1 relative to HCN2/HCN4 are attributable to higher HCN1 voltage sensors' rates and looser voltage-independent interactions between subunits in closed/open transitions. It also accounts for experimental evidence that reduction of sensors' positive charge leads to negative voltage shifts of Po curve, with little change of curve slope. HCN voltage gating thus involves two processes: voltage sensor gating and allosteric opening/closing.
超极化激活(起搏器)通道由负电压和细胞内cAMP双重门控。天然心脏f通道的动力学与霍奇金-赫胥黎(HH)门控不相符,需要封闭/开放多状态模型。我们证实,HCN通道家族成员(mHCN1、hHCN2、hHCN4)也具有不符合HH门控的特性,如S形激活和失活、激活偏离指数的固定幂次、通过预处理超极化消除激活“延迟”。先前对天然通道的研究表明,cAMP对开放概率(Po)曲线的移动作用可以用变构模型来解释,即cAMP与开放通道的结合比与封闭通道更有利。因此,我们询问超极化激活通道的不仅依赖cAMP的门控,而且依赖电压的门控是否也能用变构模型来解释。我们假设HCN通道是四聚体,每个亚基包含一个在“不情愿”和“愿意”状态之间移动的电压传感器,而电压传感器由电压独立门控,通道的封闭/开放转变通过变构发生。这些假设导致了一个包含五个开放和五个封闭通道状态的多状态方案。我们通过拟合首次激活延迟曲线和单指数时间常数曲线,然后拟合单个激活/失活轨迹来估计模型速率常数。通过简单地使用不同的速率常数集,该模型解释了所研究的所有三种HCN同工型电压门控的定性和定量方面,并允许对不同同工型的不同动力学特性进行解释。例如,HCN1相对于HCN2/HCN4更快的动力学归因于更高的HCN1电压传感器速率以及封闭/开放转变中亚基之间更松散的非电压依赖性相互作用。它还解释了实验证据,即传感器正电荷的减少导致Po曲线的负电压偏移,而曲线斜率变化很小。因此,HCN电压门控涉及两个过程:电压传感器门控和变构开放/关闭。