Parker Jeremy David Kirk, Quarmby Lynne Marie
Simon Fraser University, 8888 University Drive, Burnaby, BC, V5A 1S6, Canada.
BMC Cell Biol. 2003 Aug 20;4:11. doi: 10.1186/1471-2121-4-11.
Cilia and flagella are often lost in anticipation of mitosis or in response to stress. There are two ways that a cell can lose its flagella: resorption or deflagellation. Deflagellation involves active severing of the axoneme at the base of the flagellum; this process is defective in Chlamydomonas fa mutants. In contrast, resorption has been thought to occur as a consequence of constitutive disassembly at the tip in the absence of continued assembly, which requires intraflagellar transport (IFT). Chlamydomonas fla mutants are unable to build and maintain flagella due to defects in IFT.
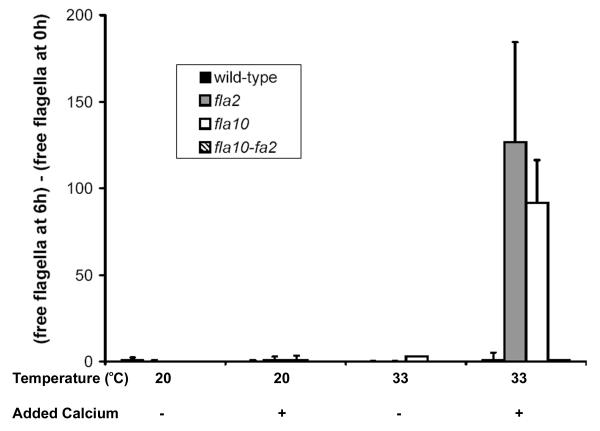
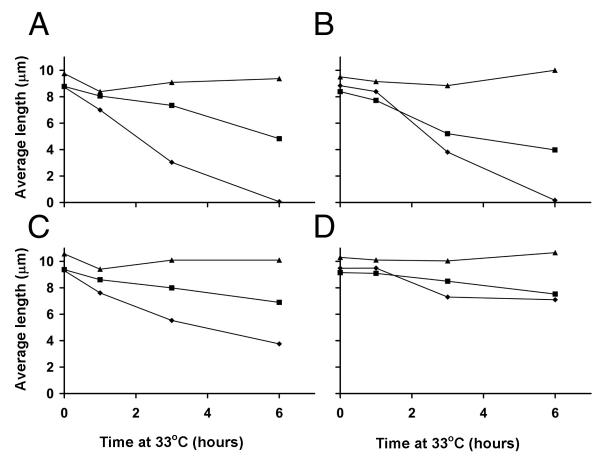
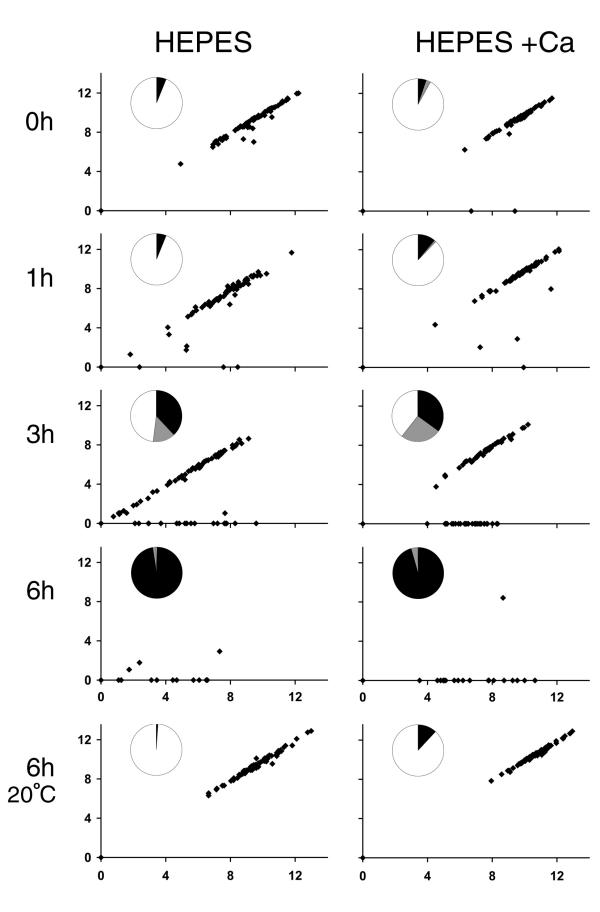
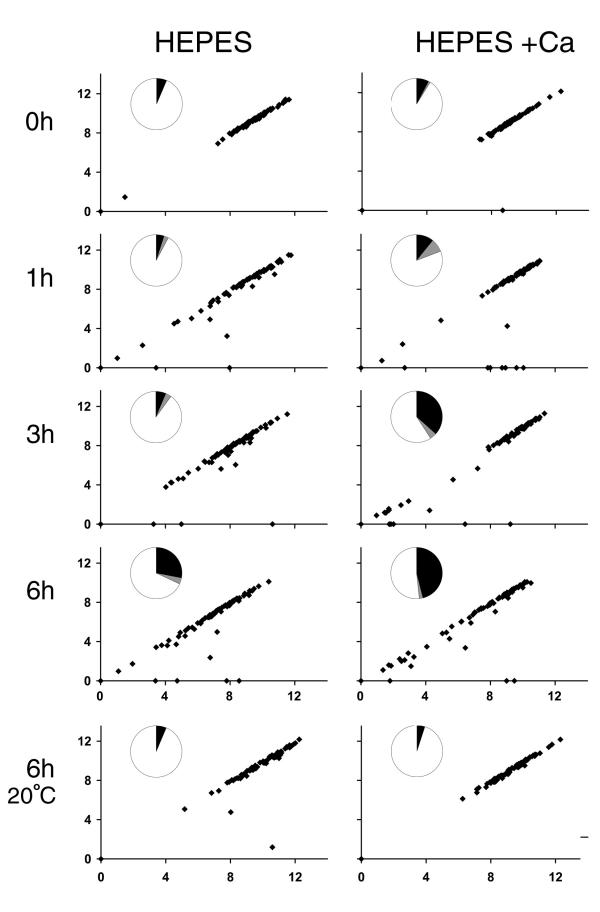
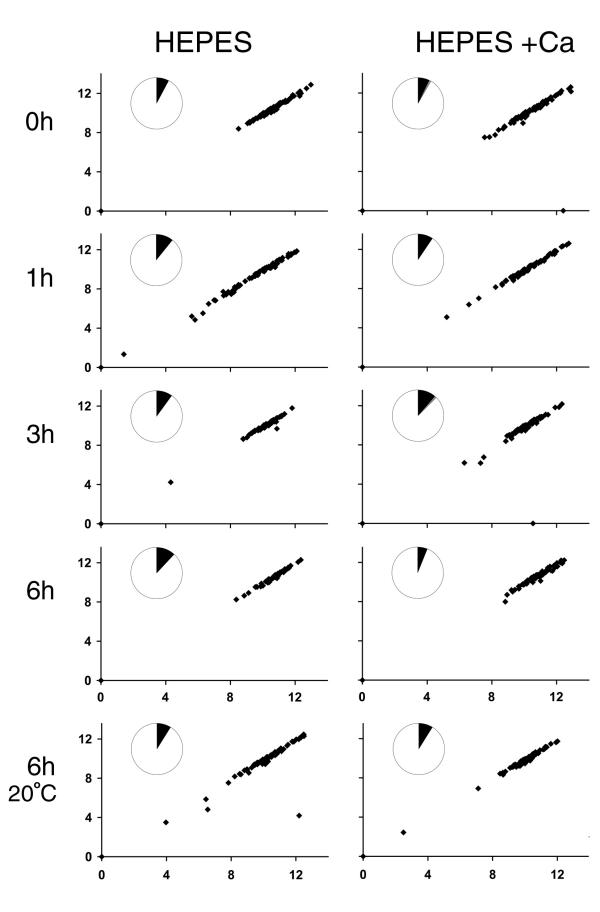
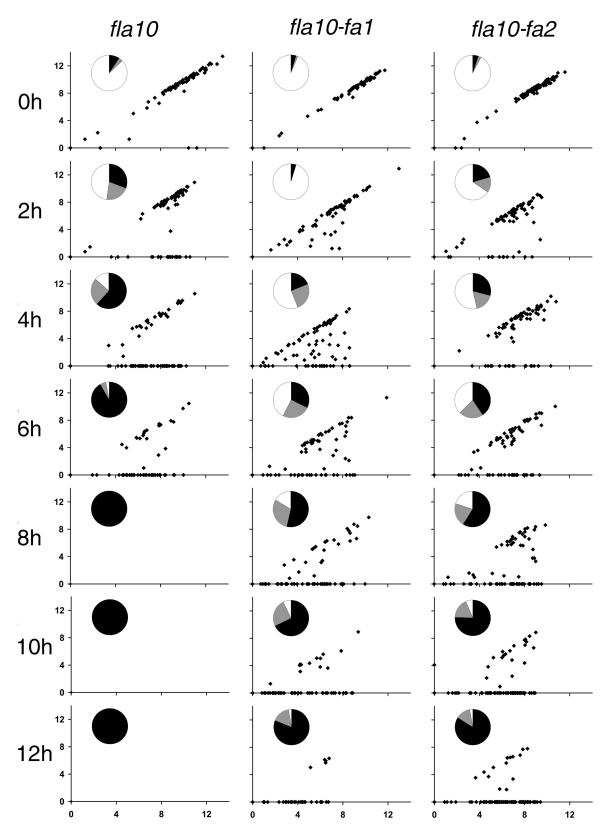
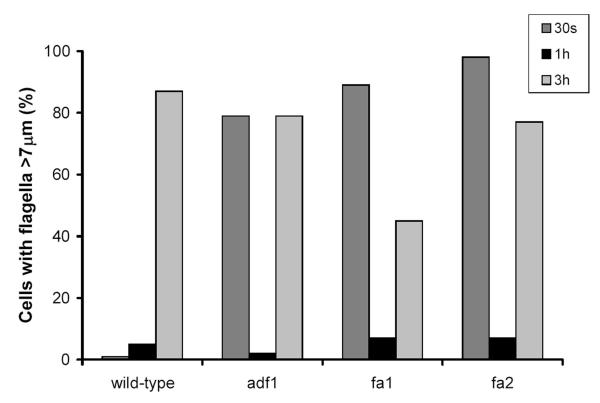
fla10 cells, which are defective in kinesin-II, the anterograde IFT motor, resorb their flagella at the restrictive temperature (33 degrees C), as previously reported. We find that in standard media containing approximately 300 microM calcium, fla10 cells lose flagella by deflagellation at 33 degrees C. This temperature-induced deflagellation of a fla mutant is not predicted by the IFT-based model for flagellar length control. Other fla mutants behave similarly, losing their flagella by deflagellation instead of resorption, if adequate calcium is available. These data suggest a new model whereby flagellar resorption involves active disassembly at the base of the flagellum via a mechanism with components in common with the severing machinery of deflagellation. As predicted by this model, we discovered that deflagellation stimuli induce resorption if deflagellation is blocked either by mutation in a FA gene or by lack of calcium. Further support for this model comes from our discovery that fla10-fa double mutants resorb their flagella more slowly than fla10 mutants.
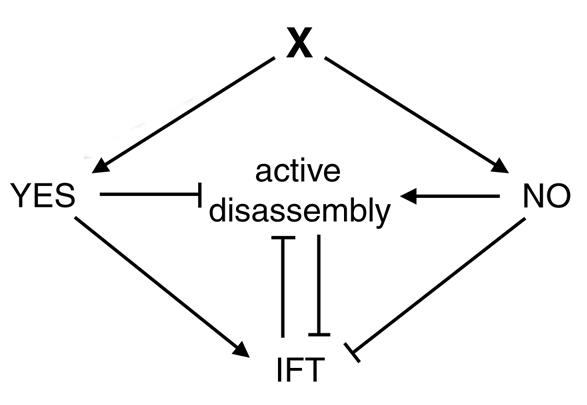
Deflagellation of the fla10 mutant at the restrictive temperature is indicative of an active disassembly signal, which can manifest as either resorption or deflagellation. We propose that when IFT is halted by either an inactivating mutation or a cellular signal, active flagellar disassembly is initiated. This active disassembly is distinct from the constitutive disassembly which plays a role in flagellar length control.
纤毛和鞭毛通常会在有丝分裂预期时或应激反应中丢失。细胞失去鞭毛有两种方式:吸收或去鞭毛作用。去鞭毛作用涉及在鞭毛基部对轴丝进行主动切断;此过程在衣藻fa突变体中存在缺陷。相比之下,吸收被认为是在缺乏持续组装(这需要鞭毛内运输,即IFT)的情况下,由于鞭毛顶端的组成性解聚而发生的。衣藻fla突变体由于IFT缺陷而无法构建和维持鞭毛。
fla10细胞在驱动IFT正向运输的动力蛋白II方面存在缺陷,如先前报道,它们在限制温度(33摄氏度)下会吸收其鞭毛。我们发现,在含有约300微摩尔钙的标准培养基中,fla10细胞在33摄氏度时通过去鞭毛作用失去鞭毛。基于IFT的鞭毛长度控制模型无法预测fla突变体的这种温度诱导的去鞭毛作用。如果有足够的钙,其他fla突变体也有类似表现,通过去鞭毛作用而非吸收失去鞭毛。这些数据提示了一种新模型,即鞭毛吸收涉及通过一种与去鞭毛切断机制有共同成分的机制在鞭毛基部进行主动解聚。正如该模型所预测的,我们发现如果去鞭毛作用因FA基因突变或缺乏钙而受阻,去鞭毛刺激会诱导吸收。对该模型的进一步支持来自我们的发现,即fla10 - fa双突变体比fla10突变体吸收鞭毛的速度更慢。
fla10突变体在限制温度下的去鞭毛作用表明存在一个主动解聚信号,该信号可表现为吸收或去鞭毛作用。我们提出,当IFT因失活突变或细胞信号而停止时,会启动主动的鞭毛解聚。这种主动解聚不同于在鞭毛长度控制中起作用的组成性解聚。