De Rijcke M, Seneca S, Punyammalee B, Glansdorff N, Crabeel M
Erfelijkheidsleer en Microbiologie, Vrije Universiteit Brussel, Belgium.
Mol Cell Biol. 1992 Jan;12(1):68-81. doi: 10.1128/mcb.12.1.68-81.1992.
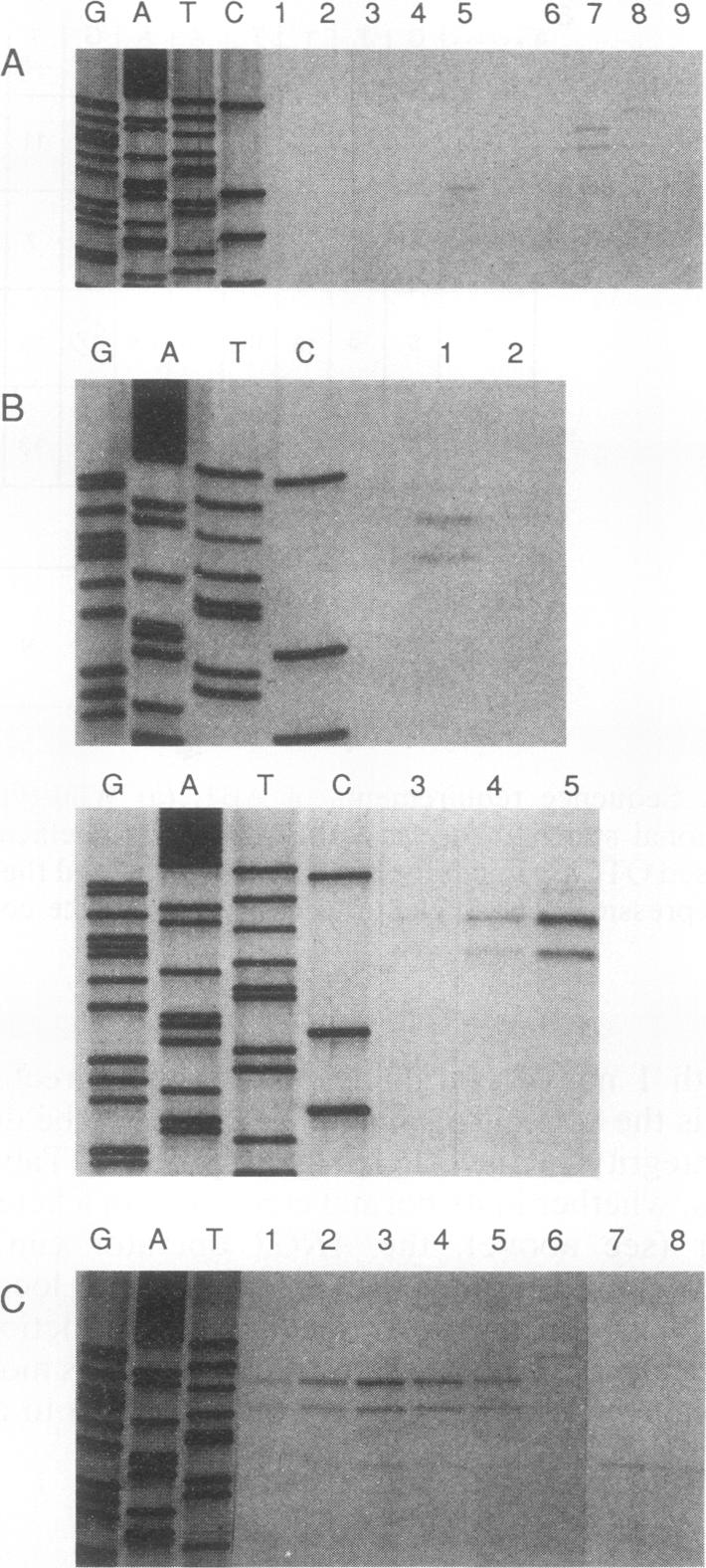
We have determined the sequences and positions of the cis elements required for proper functioning of the ARG3 promoter and proper arginine-specific control. A TATA box located 100 nucleotides upstream of the transcription start was shown to be essential for ARG3 transcription. Two sequences involved in normal arginine-mediated repression lie immediately downstream of the TATA box: an essential one (arginine box 1 [AB1]) and a secondary one (arginine box 2 [AB2]). AB1 was defined by saturation mutagenesis and is an asymmetrical sequence. A stringently required CGPu motif in AB1 is conserved in all known target sites of C6 zinc cluster DNA-binding proteins, leading us to propose that AB1 is the binding site of ARGRII, another member of the C6 family. The palindromic AB2 sequence is suggested, on the basis of published data, to be the binding site of ARGRI, possibly in heterodimerization with MCM1. AB2 and AB1 correspond respectively to the 5' and 3' halves of two adjacent similar sequences of 29 bp that appear to constitute tandem operators. Indeed, mutations increasing the similarity of the other halves with AB1 and AB2 cause hyperrepression. To mediate repression, the operator must be located close to the transcription initiation region. It remains functional if the TATA box is moved downstream of it but becomes inoperative in repression when displaced to a far-upstream position where it mediates an arginine and ARGR-dependent induction of gene expression. The ability of the ARG3 operator to act either as an operator or as an upstream activator sequence, depending on its location, and the functional organization of the anabolic and catabolic arginine genes suggest a simple model for arginine regulation in which an activator complex can turn into a repressor when able to interfere sterically with the process of transcription initiation.
我们已经确定了ARG3启动子正常发挥功能以及精氨酸特异性调控所需的顺式元件的序列和位置。位于转录起始点上游100个核苷酸处的TATA盒对于ARG3转录至关重要。参与正常精氨酸介导的阻遏作用的两个序列紧邻TATA盒下游:一个是必需序列(精氨酸盒1 [AB1]),另一个是次要序列(精氨酸盒2 [AB2])。AB1通过饱和诱变确定,是一个不对称序列。AB1中一个严格必需的CGPu基序在所有已知的C6锌簇DNA结合蛋白的靶位点中保守,这使我们提出AB1是C6家族的另一个成员ARGRII的结合位点。根据已发表的数据,回文AB2序列被认为是ARGRI的结合位点,可能与MCM1形成异二聚体。AB2和AB1分别对应于两个相邻的29 bp相似序列的5'和3'半段,这两个序列似乎构成串联操纵子。实际上,增加另一半与AB1和AB2相似性的突变会导致超阻遏。为了介导阻遏,操纵子必须位于靠近转录起始区域的位置。如果TATA盒移至其下游,它仍然具有功能,但当移至远上游位置时,它在阻遏中变得不起作用,在该位置它介导精氨酸和ARGR依赖的基因表达诱导。ARG3操纵子根据其位置既可以作为操纵子也可以作为上游激活序列发挥作用的能力,以及合成代谢和分解代谢精氨酸基因的功能组织,提示了一个简单的精氨酸调控模型,即当激活复合物能够在空间上干扰转录起始过程时,它可以转变为阻遏物。