Koumandou V Lila, Dacks Joel B, Coulson Richard M R, Field Mark C
Department of Pathology, University of Cambridge, Tennis Court Road, Cambridge, CB2 1QP, UK.
BMC Evol Biol. 2007 Feb 23;7:29. doi: 10.1186/1471-2148-7-29.
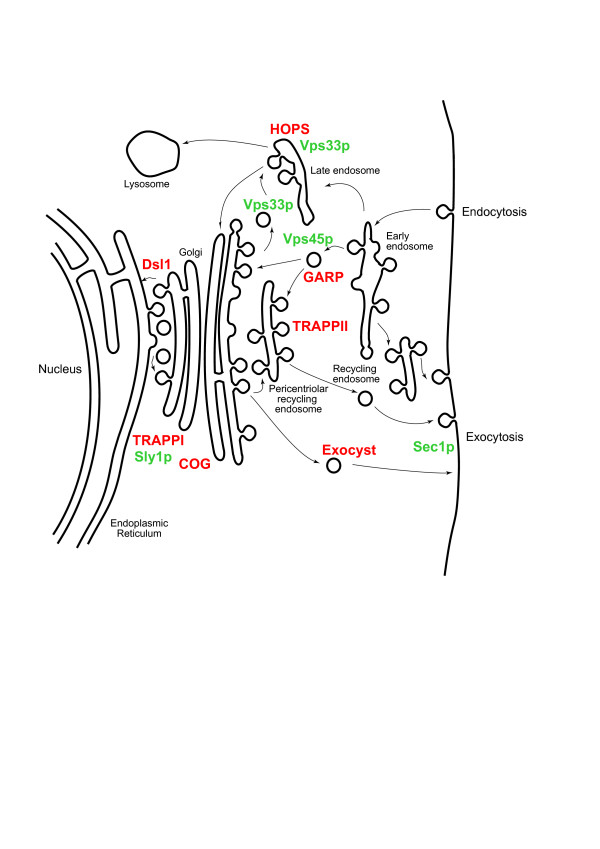
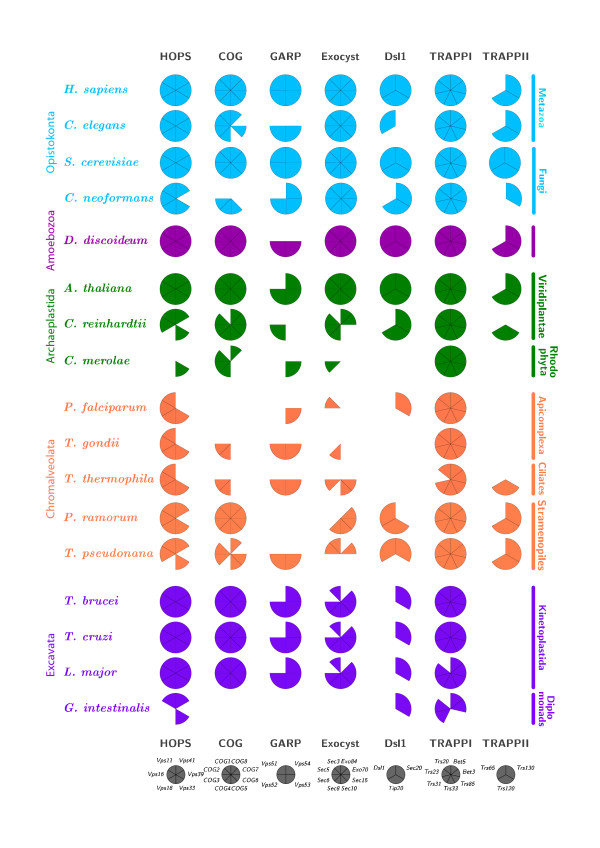
In membrane trafficking, the mechanisms ensuring vesicle fusion specificity remain to be fully elucidated. Early models proposed that specificity was encoded entirely by SNARE proteins; more recent models include contributions from Rab proteins, Syntaxin-binding (SM) proteins and tethering factors. Most information on membrane trafficking derives from an evolutionarily narrow sampling of model organisms. However, considering factors from a wider diversity of eukaryotes can provide both functional information on core systems and insight into the evolutionary history of the trafficking machinery. For example, the major Qa/syntaxin SNARE families are present in most eukaryotic genomes and likely each evolved via gene duplication from a single ancestral syntaxin before the existing eukaryotic groups diversified. This pattern is also likely for Rabs and various other components of the membrane trafficking machinery.
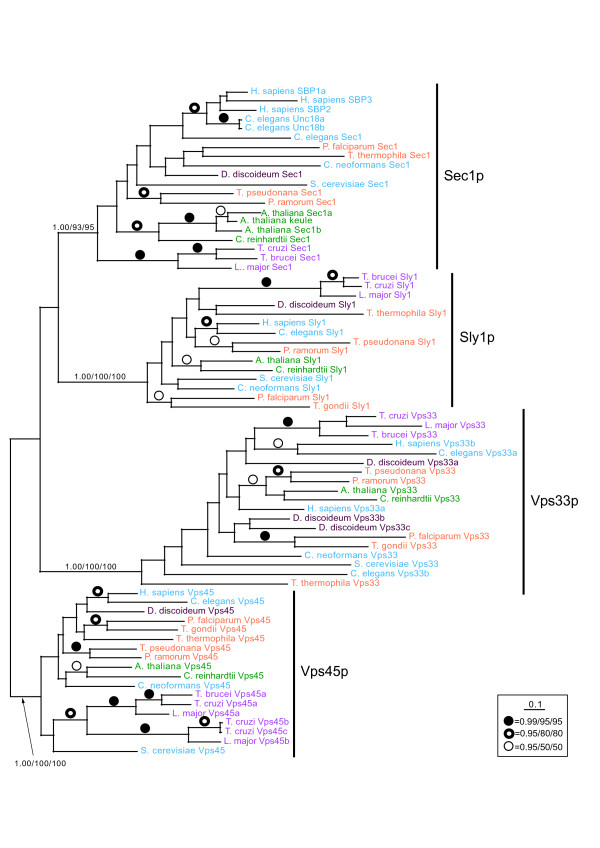
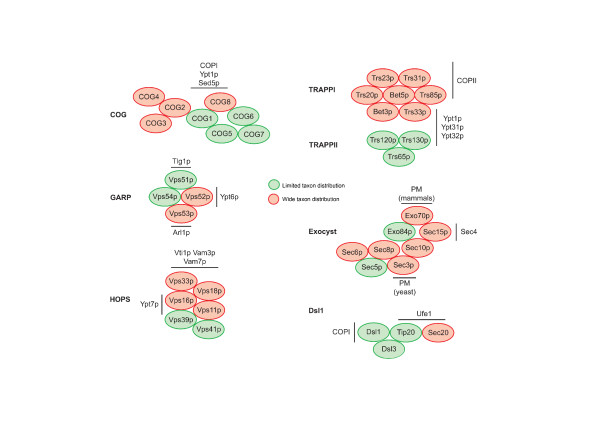
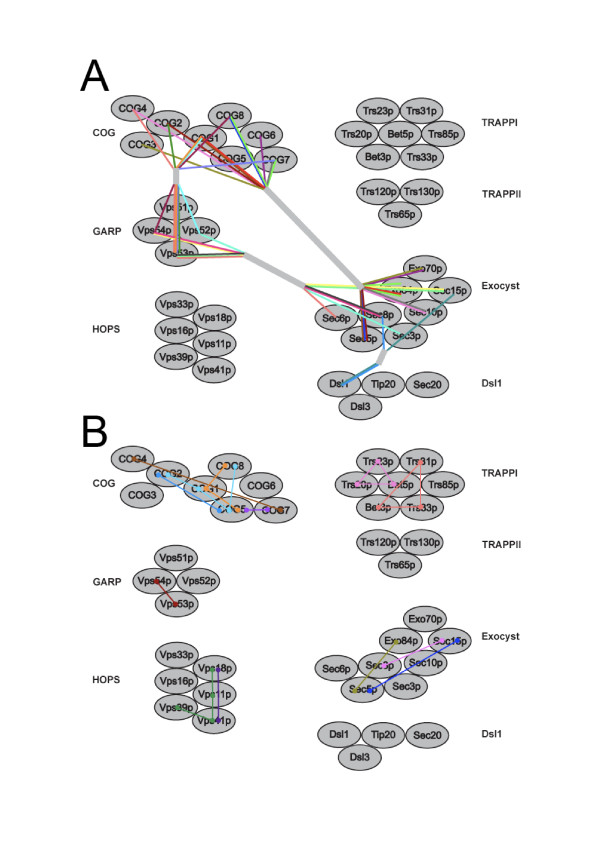
We performed comparative genomic and phylogenetic analyses, when relevant, on the SM proteins and components of the tethering complexes, both thought to contribute to vesicle fusion specificity. Despite evidence suggestive of secondary losses amongst many lineages, the tethering complexes are well represented across the eukaryotes, suggesting an origin predating the radiation of eukaryotic lineages. Further, whilst we detect distant sequence relations between GARP, COG, exocyst and DSL1 components, these similarities most likely reflect convergent evolution of similar secondary structural elements. No similarity is found between the TRAPP and HOPS complexes and the other tethering factors. Overall, our data favour independent origins for the various tethering complexes. The taxa examined possess at least one homologue of each of the four SM protein families; since the four monophyletic families each encompass a wide diversity of eukaryotes, the SM protein families very likely evolved before the last common eukaryotic ancestor (LCEA).
These data further support a highly complex LCEA and indicate that the basic architecture of the trafficking system is remarkably conserved and ancient, with the SM proteins and tethering factors having originated very early in eukaryotic evolution. However, the independent origin of the tethering complexes suggests a novel pattern for increasing complexity in the membrane trafficking system, in addition to the pattern of paralogous machinery elaboration seen thus far.
在膜泡运输过程中,确保囊泡融合特异性的机制仍有待充分阐明。早期模型认为特异性完全由SNARE蛋白编码;最近的模型则包括Rab蛋白、Syntaxin结合(SM)蛋白和拴系因子的作用。关于膜泡运输的大多数信息来自进化上有限的模式生物样本。然而,考虑来自更广泛多样的真核生物的因素,既可以提供关于核心系统的功能信息,也能深入了解运输机制的进化历史。例如,大多数真核生物基因组中都存在主要的Qa/Syntaxin SNARE家族,并且在现有真核生物类群分化之前,每个家族可能都通过基因复制从单一的祖先Syntaxin进化而来。Rabs和膜泡运输机制的各种其他成分可能也是这种模式。
我们在相关情况下对SM蛋白和拴系复合体的成分进行了比较基因组学和系统发育分析,二者都被认为对囊泡融合特异性有贡献。尽管有证据表明许多谱系中存在次生丢失,但拴系复合体在整个真核生物中都有很好的代表性,这表明其起源早于真核生物谱系的辐射。此外,虽然我们检测到GARP、COG、外泌体和DSL1成分之间存在远缘序列关系,但这些相似性很可能反映了相似二级结构元件的趋同进化。TRAPP和HOPS复合体与其他拴系因子之间未发现相似性。总体而言,我们的数据支持各种拴系复合体的独立起源。所研究的分类群至少拥有四个SM蛋白家族中每个家族的一个同源物;由于这四个单系家族各自涵盖了广泛多样的真核生物,因此SM蛋白家族很可能在最后一个共同的真核祖先(LCEA)之前就已进化。
这些数据进一步支持了一个高度复杂的LCEA,并表明运输系统基本结构显著保守且古老,SM蛋白和拴系因子在真核生物进化早期就已起源。然而,拴系复合体的独立起源表明,除了迄今为止所见的旁系同源机制细化模式外,膜泡运输系统复杂性增加还有一种新模式。