Teichman Shlomit, Kanner Baruch I
Department of Biochemistry, Hebrew University Hadassah Medical School, Jerusalem 91120, Israel.
J Gen Physiol. 2007 Jun;129(6):527-39. doi: 10.1085/jgp.200609707.
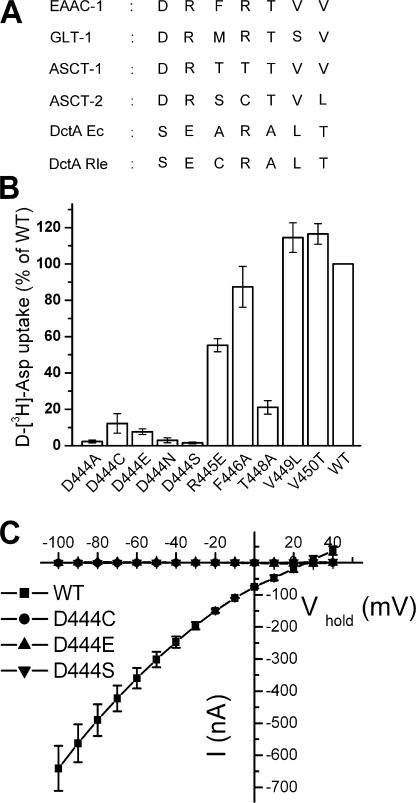
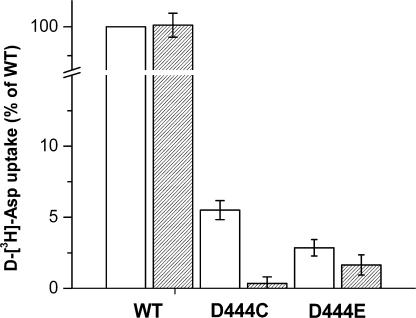
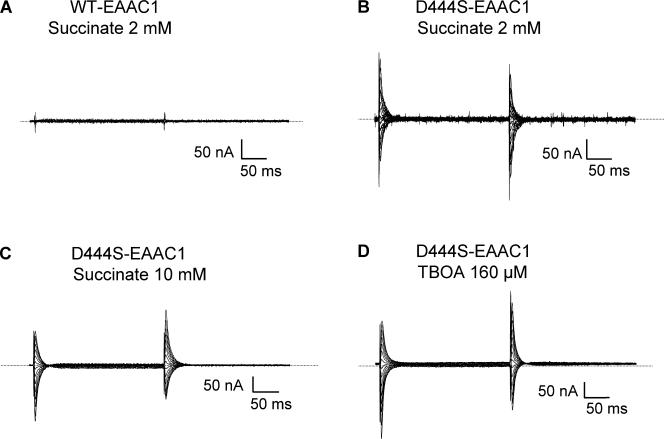
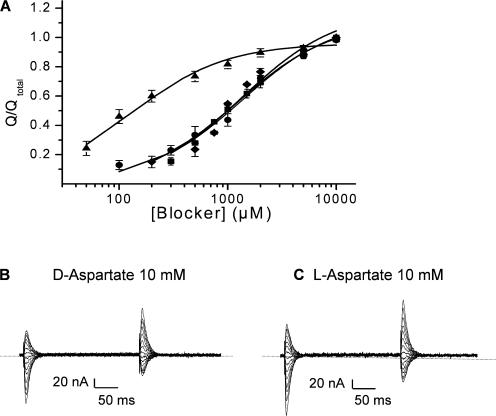
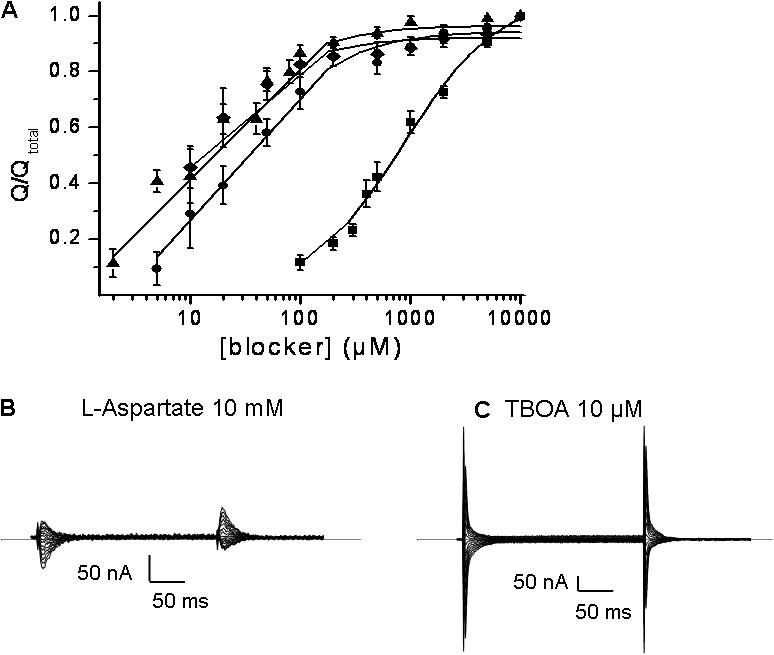
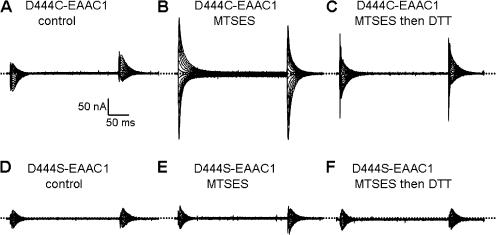
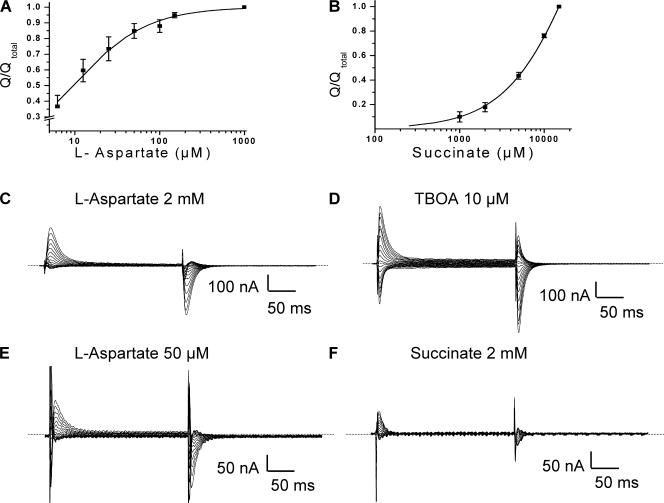
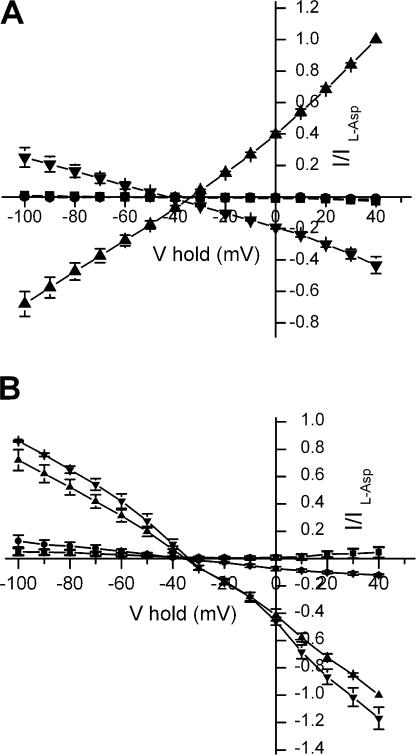
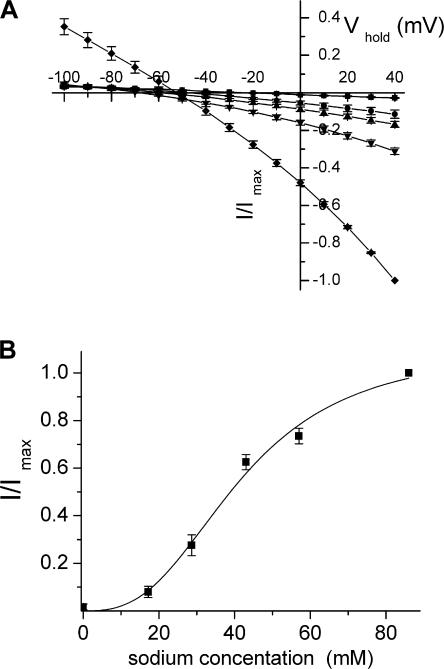
In the central nervous system, electrogenic sodium- and potassium-coupled glutamate transporters terminate the synaptic actions of this neurotransmitter. In contrast to acidic amino acids, dicarboxylic acids are not recognized by glutamate transporters, but the related bacterial DctA transporters are capable of transporting succinate and other dicarboxylic acids. Transmembrane domain 8 contains several residues that differ between these two types of transporters. One of these, aspartate-444 of the neuronal glutamate transporter EAAC1, is conserved in glutamate transporters, but a serine residue occupies this position in DctA transporters. When aspartate-444 is mutated to serine, cysteine, alanine, or even to glutamate, uptake of D-[(3)H]-aspartate as well as the inwardly rectifying steady-state currents induced by acidic amino acids is impaired. Even though succinate was not capable of inducing any steady-state transport currents, the dicarboxylic acid inhibited the sodium-dependent transient currents by the mutants with a neutral substitution at position 444. In the neutral substitution mutants inhibition of the transients was also observed with acidic amino acids. In the D444E mutant, acidic amino acids were potent inhibitors of the transient currents, whereas the apparent affinity for succinate was lower by at least three orders of magnitude. Even though L-aspartate could bind to D444E with a high apparent affinity, this binding resulted in inhibition rather than stimulation of the uncoupled anion conductance. Thus, a carboxylic acid-containing side chain at position 444 prevents the interaction of glutamate transporters with succinate, and the presence of aspartate itself at this position is crucial for productive substrate binding compatible with substrate translocation.
在中枢神经系统中,电生性钠钾偶联谷氨酸转运体终止了这种神经递质的突触作用。与酸性氨基酸不同,二羧酸不被谷氨酸转运体识别,但相关的细菌DctA转运体能够转运琥珀酸和其他二羧酸。跨膜结构域8包含这两种转运体之间不同的几个残基。其中之一是神经元谷氨酸转运体EAAC1的天冬氨酸-444,它在谷氨酸转运体中保守,但在DctA转运体中此位置为丝氨酸残基。当将天冬氨酸-444突变为丝氨酸、半胱氨酸、丙氨酸,甚至谷氨酸时,D-[(3)H]-天冬氨酸的摄取以及酸性氨基酸诱导的内向整流稳态电流均受损。尽管琥珀酸不能诱导任何稳态转运电流,但这种二羧酸抑制了444位为中性取代的突变体的钠依赖性瞬态电流。在中性取代突变体中,酸性氨基酸也能抑制瞬态电流。在D444E突变体中,酸性氨基酸是瞬态电流的有效抑制剂,而对琥珀酸的表观亲和力至少低三个数量级。尽管L-天冬氨酸能以高表观亲和力与D444E结合,但这种结合导致未偶联阴离子电导的抑制而非刺激。因此,444位含羧酸的侧链阻止了谷氨酸转运体与琥珀酸的相互作用,此位置天冬氨酸本身的存在对于与底物转运相容的有效底物结合至关重要。