Departamento de Ingeniería Celular y Biocatálisis, Instituto de Biotecnología, Universidad Nacional Autónoma de México, Av, Universidad 2001, Cuernavaca, Morelos, México.
Microb Cell Fact. 2007 Sep 19;6:30. doi: 10.1186/1475-2859-6-30.
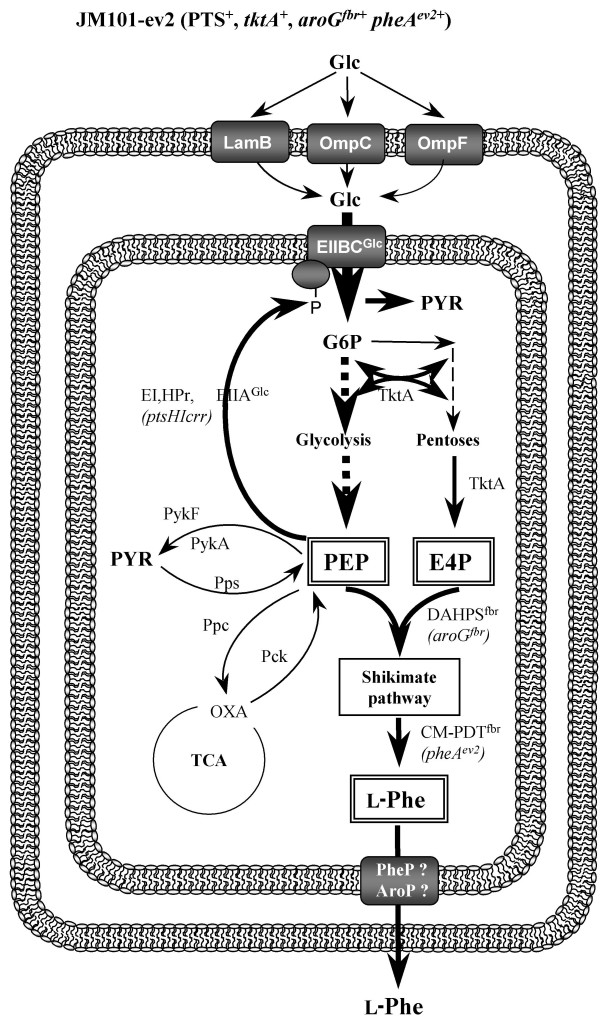
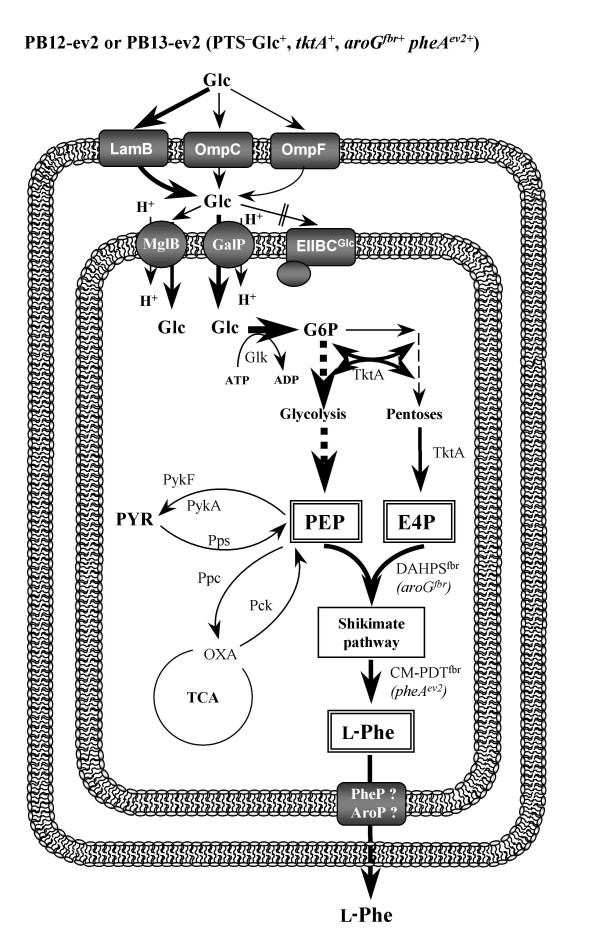
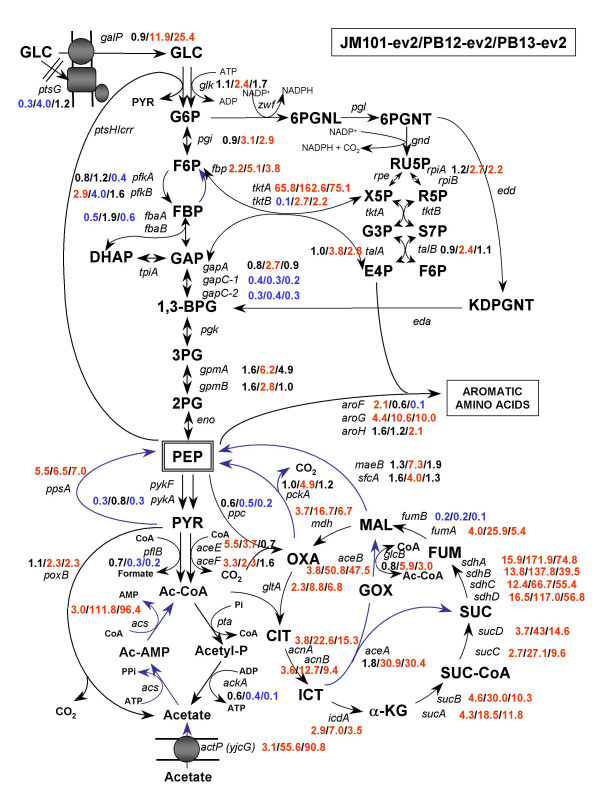
The rational design of L-phenylalanine (L-Phe) overproducing microorganisms has been successfully achieved by combining different genetic strategies such as inactivation of the phosphoenolpyruvate: phosphotransferase transport system (PTS) and overexpression of key genes (DAHP synthase, transketolase and chorismate mutase-prephenate dehydratase), reaching yields of 0.33 (g-Phe/g-Glc), which correspond to 60% of theoretical maximum. Although genetic modifications introduced into the cell for the generation of overproducing organisms are specifically targeted to a particular pathway, these can trigger unexpected transcriptional responses of several genes. In the current work, metabolic transcription analysis (MTA) of both L-Phe overproducing and non-engineered strains using Real-Time PCR was performed, allowing the detection of transcriptional responses to PTS deletion and plasmid presence of genes related to central carbon metabolism. This MTA included 86 genes encoding enzymes of glycolysis, gluconeogenesis, pentoses phosphate, tricarboxylic acid cycle, fermentative and aromatic amino acid pathways. In addition, 30 genes encoding regulatory proteins and transporters for aromatic compounds and carbohydrates were also analyzed.
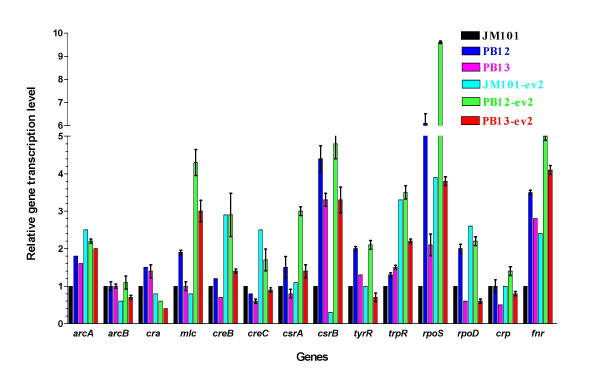
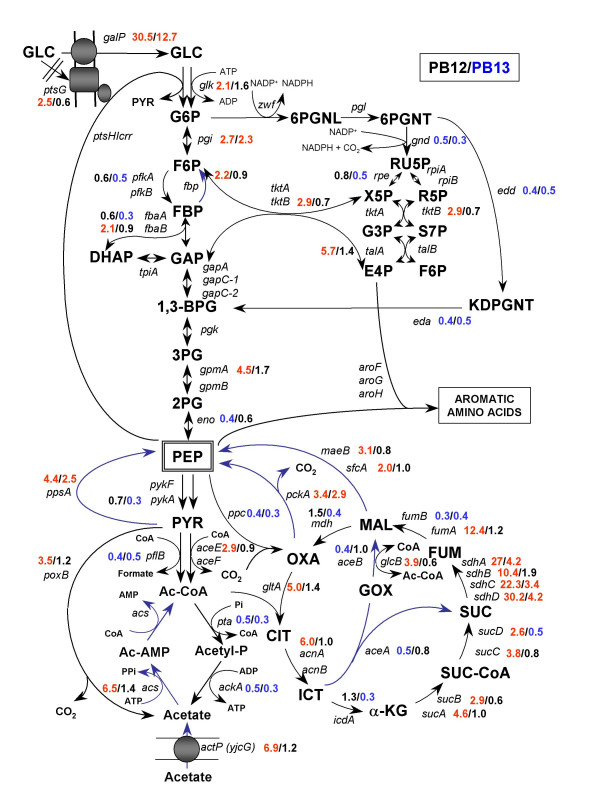
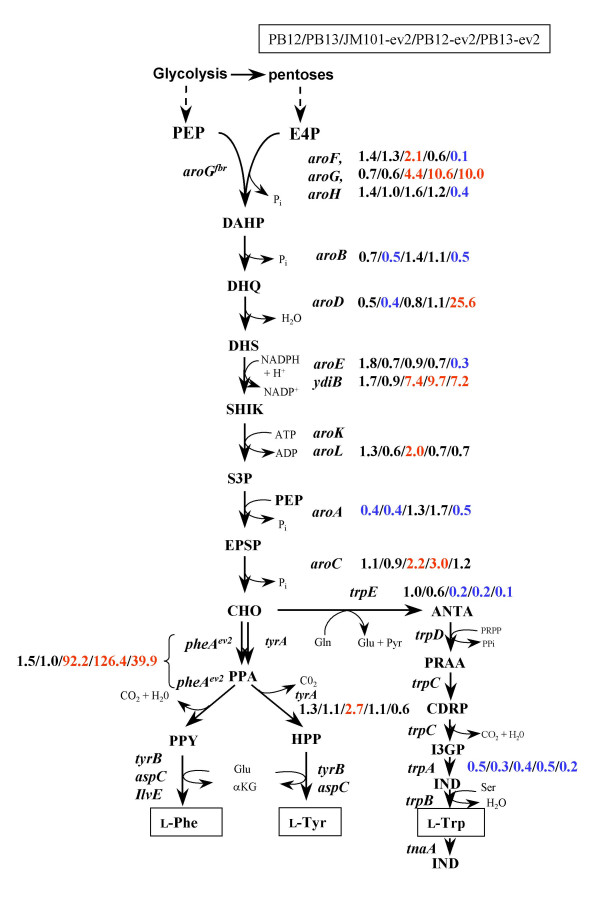
MTA revealed that a set of genes encoding carbohydrate transporters (galP, mglB), gluconeogenic (ppsA, pckA) and fermentative enzymes (ldhA) were significantly induced, while some others were down-regulated such as ppc, pflB, pta and ackA, as a consequence of PTS inactivation. One of the most relevant findings was the coordinated up-regulation of several genes that are exclusively gluconeogenic (fbp, ppsA, pckA, maeB, sfcA, and glyoxylate shunt) in the best PTS- L-Phe overproducing strain (PB12-ev2). Furthermore, it was noticeable that most of the TCA genes showed a strong up-regulation in the presence of multicopy plasmids by an unknown mechanism. A group of genes exhibited transcriptional responses to both PTS inactivation and the presence of plasmids. For instance, acs-ackA, sucABCD, and sdhABCD operons were up-regulated in PB12 (PTS mutant that carries an arcB- mutation). The induction of these operons was further increased by the presence of plasmids in PB12-ev2. Some genes involved in the shikimate and specific aromatic amino acid pathways showed down-regulation in the L-Phe overproducing strains, might cause possible metabolic limitations in the shikimate pathway.
The identification of potential rate-limiting steps and the detection of transcriptional responses in overproducing microorganisms may suggest "reverse engineering" strategies for the further improvement of L-Phe production strains.
通过结合不同的遗传策略,如磷酸烯醇丙酮酸:磷酸转移酶运输系统(PTS)的失活和关键基因(DAHP 合酶、转酮醇酶和磷酸苯丙氨酸脱水酶)的过表达,已经成功地实现了 L-苯丙氨酸(L-Phe)的理性设计。产量达到 0.33(g-Phe/g-Glc),相当于理论最大值的 60%。尽管为产生高产微生物而引入细胞的遗传修饰专门针对特定途径,但这些修饰可能会引发几个基因的意外转录反应。在目前的工作中,使用实时 PCR 对 L-Phe 高产菌株和非工程菌株进行了代谢转录分析(MTA),从而可以检测到 PTS 缺失和与中心碳代谢相关的基因质粒存在的转录反应。该 MTA 包括 86 个编码糖酵解、糖异生、戊糖磷酸、三羧酸循环、发酵和芳香族氨基酸途径的酶的基因。此外,还分析了 30 个编码芳香族化合物和碳水化合物的调节蛋白和转运蛋白的基因。
MTA 显示,一组编码碳水化合物转运蛋白(galP、mglB)、糖异生(ppsA、pckA)和发酵酶(ldhA)的基因被显著诱导,而一些其他基因被下调,如 ppc、pflB、pta 和 ackA,这是 PTS 失活的结果。最相关的发现之一是,在最佳 PTS-L-Phe 高产菌株(PB12-ev2)中,一组专门用于糖异生的基因(fbp、ppsA、pckA、maeB、sfcA 和乙醛酸支路)被协调上调。此外,值得注意的是,大多数 TCA 基因通过未知机制在多拷贝质粒存在下表现出强烈的上调。一组基因表现出对 PTS 失活和质粒存在的转录反应。例如,acs-ackA、sucABCD 和 sdhABCD 操纵子在 PTS 突变(携带 arcB-突变的 PB12)中被上调。在 PB12-ev2 中存在质粒时,这些操纵子的诱导进一步增加。一些参与莽草酸和特定芳香族氨基酸途径的基因在 L-Phe 高产菌株中下调,可能导致莽草酸途径中存在潜在的代谢限制。
鉴定高产微生物中的潜在限速步骤和检测转录反应可能为进一步提高 L-Phe 生产菌株提出“反向工程”策略。