Jugdaohsingh Ravin, Calomme Mario R, Robinson Karen, Nielsen Forrest, Anderson Simon H C, D'Haese Patrick, Geusens Piet, Loveridge Nigel, Thompson Richard P H, Powell Jonathan J
MRC Human Nutrition Research, Elsie Widdowson Laboratory, Fulbourn Road, Cambridge CB1 9NL, UK.
Bone. 2008 Sep;43(3):596-606. doi: 10.1016/j.bone.2008.04.014. Epub 2008 May 2.
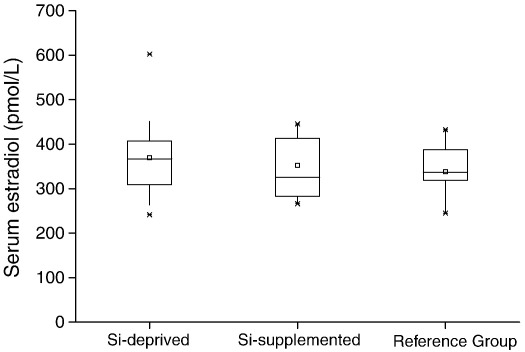
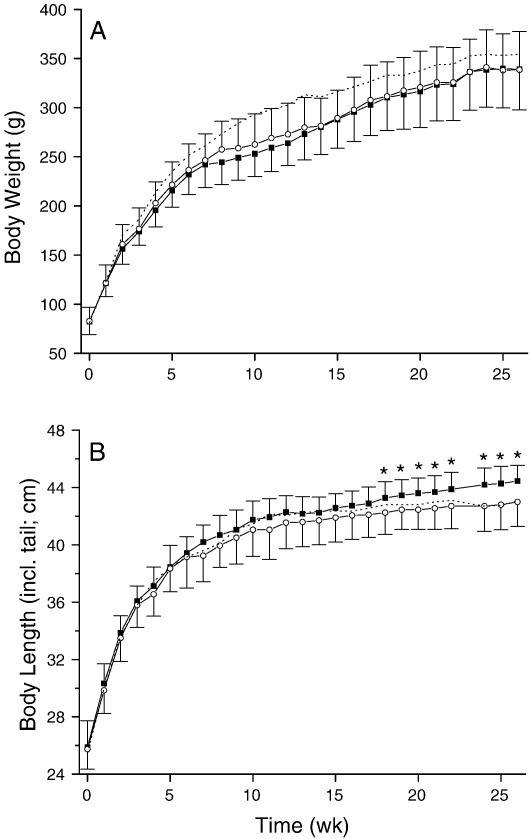
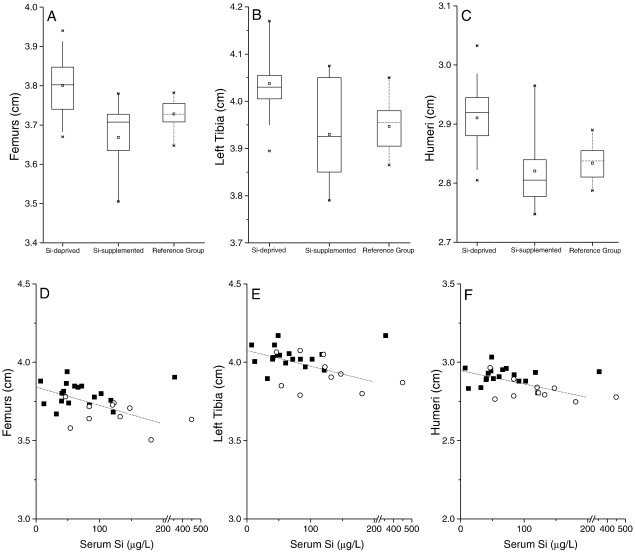
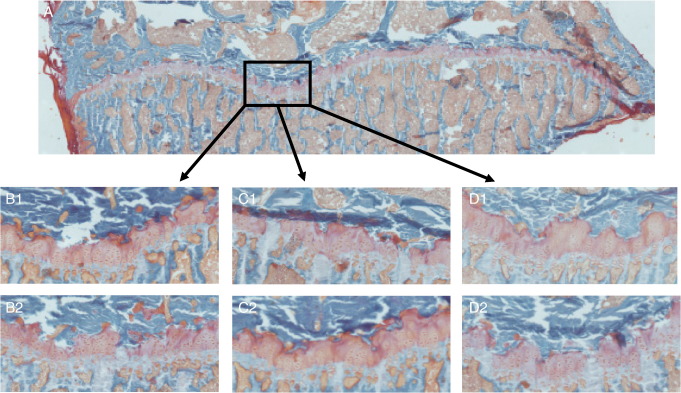
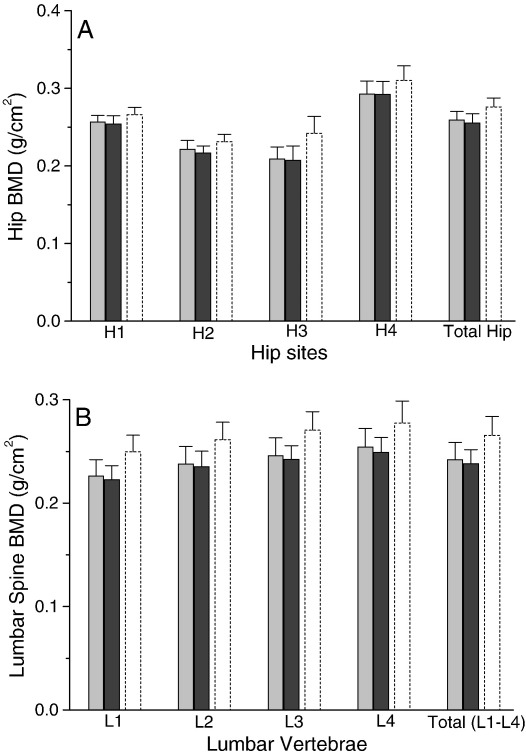
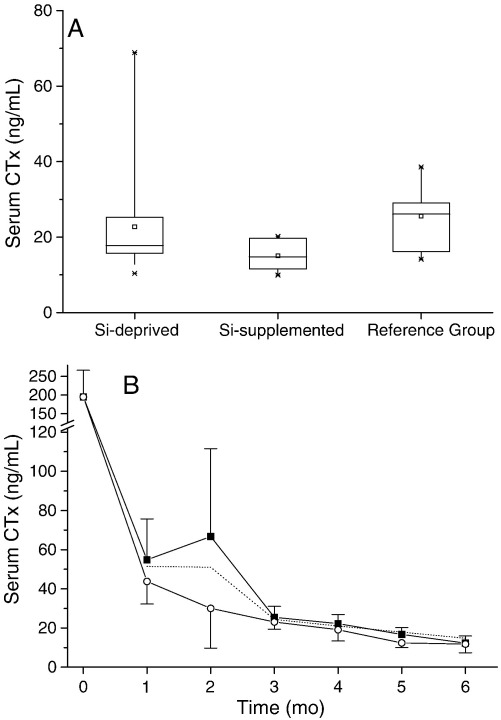
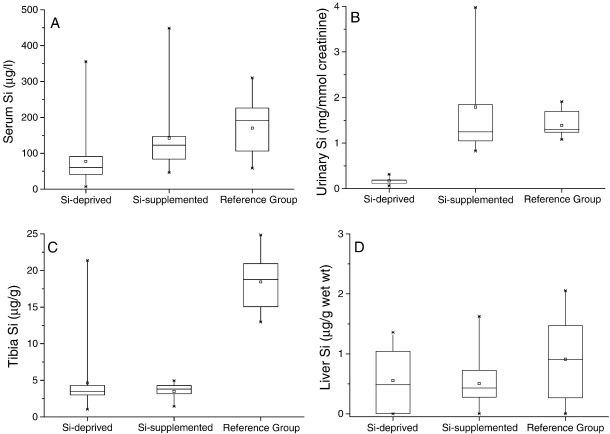
Silicon-deficiency studies in growing animals in the early 1970s reported stunted growth and profound defects in bone and other connective tissues. However, more recent attempts to replicate these findings have found mild alterations in bone metabolism without any adverse health effects. Thus the biological role of silicon remains unknown. Using a specifically formulated silicon-depleted diet and modern methods for silicon analysis and assessment of skeletal development, we undertook, through international collaboration between silicon researchers, an extensive study of long-term silicon depletion on skeletal development in an animal. 21-day old female Sprague-Dawley rats (n=20) were fed a silicon-depleted diet (3.2 microg Si/g feed) for 26 weeks and their growth and skeletal development were compared with identical rats (n=10) on the same diet but with silicon added as Si(OH)(4) to their drinking water (53.2 microg Si/g water); total silicon intakes were 24 times different. A third group of rats, receiving a standard rodent stock feed (322 microg Si/g feed) and tap water (5 microg Si/g water), served as a reference group for optimal growth. A series of anthropometric and bone quality measures were undertaken during and following the study. Fasting serum silicon concentrations and especially urinary silicon excretion were significantly lower in the silicon-deprived group compared to the supplemented group (P=0.03 and 0.004, respectively). Tibia and soft-tissue silicon contents did not differ between the two groups, but tibia silicon levels were significantly lower compared to the reference group (P<0.0001). Outward adverse health effects were not observed in the silicon-deprived group. However, body lengths from week 18 onwards (P<0.05) and bone lengths at necropsy (P</=0.002) were longer in this group. Moreover, these measures correlated inversely with serum silicon concentrations (P</=0.02). A reduction in bone growth plate thickness and an apparent increase in chondrocyte density were also observed in the silicon-deprived animals. No other differences were observed between the two groups, except for tibia phosphorus concentrations, which were lower in the silicon-deprived animals (P=0.0003). Thus in this study we were unable to reproduce the profound deficiency state reported in rats and chicks in the early 1970s. Indeed, although silicon intake and circulating fasting serum levels differed between the silicon-deprived and silicon-supplemented animals, tibia and soft-tissue levels did not and may explain the lack of difference in bone quality and bone markers (except serum CTx) between these two groups. Markedly higher tibia silicon levels in the reference group and nutritional differences between the formulated low-Si and reference diets suggest that one or more co-factors may be absent from the low-Si diet that affect silicon incorporation into bone. However, evidence for urinary silicon conservation (to maintain tissue levels), changes in bone/body lengths, bone calcium:phosphorus ratio and differences at the growth plate with silicon deprivation are all novel and deserve further study. These results suggest that rats actively maintain body silicon levels via urinary conservation, but the low circulating serum silicon levels during silicon deficiency result in inhibition of growth plate closure and increased longitudinal growth. Silicon-responsive genes and Si transporters are being investigated in the kidneys of these rats.
20世纪70年代初对生长中的动物进行的缺硅研究报告称,动物生长发育迟缓,骨骼和其他结缔组织存在严重缺陷。然而,最近重复这些研究结果的尝试发现,骨骼代谢仅有轻微改变,并未对健康产生任何不良影响。因此,硅的生物学作用仍然未知。我们通过硅研究人员之间的国际合作,采用专门配制的低硅饮食以及现代硅分析方法和骨骼发育评估方法,对动物长期缺硅对骨骼发育的影响进行了广泛研究。将21日龄雌性斯普拉格-道利大鼠(n = 20)喂食低硅饮食(3.2微克硅/克饲料)26周,并将其生长和骨骼发育情况与相同饮食但在饮用水中添加了Si(OH)₄形式硅(53.2微克硅/克水)的同窝大鼠(n = 10)进行比较;总硅摄入量相差24倍。第三组大鼠喂食标准啮齿动物饲料(322微克硅/克饲料)和自来水(5微克硅/克水),作为最佳生长的参考组。在研究期间及之后进行了一系列人体测量和骨质量测量。与补充硅的组相比,缺硅组的空腹血清硅浓度,尤其是尿硅排泄量显著降低(分别为P = 0.03和0.004)。两组之间胫骨和软组织的硅含量没有差异,但与参考组相比,胫骨硅水平显著降低(P < 0.0001)。在缺硅组未观察到明显的不良健康影响。然而,该组从第18周起的体长(P < 0.05)和尸检时的骨长(P≤0.002)更长。此外,这些测量结果与血清硅浓度呈负相关(P≤0.02)。在缺硅动物中还观察到骨生长板厚度减小和软骨细胞密度明显增加。除了缺硅动物的胫骨磷浓度较低(P = 0.0003)外,两组之间未观察到其他差异。因此,在本研究中,我们无法重现20世纪70年代初在大鼠和雏鸡中报告的严重缺乏状态。事实上,尽管缺硅和补充硅的动物之间硅摄入量和循环空腹血清水平不同,但胫骨和软组织水平并无差异,这可能解释了两组之间骨质量和骨标志物(血清I型胶原交联C端肽除外)缺乏差异的原因。参考组胫骨硅水平明显更高,以及配制的低硅饮食和参考饮食之间的营养差异表明,低硅饮食中可能缺乏一种或多种影响硅掺入骨骼的辅助因子。然而,尿硅保留(以维持组织水平)、骨/体长变化、骨钙:磷比值以及缺硅时生长板差异的证据都是新发现,值得进一步研究。这些结果表明,大鼠通过尿保留积极维持体内硅水平,但缺硅期间循环血清硅水平较低会导致生长板闭合受抑制,纵向生长增加。目前正在对这些大鼠的肾脏中的硅反应基因和硅转运体进行研究。