Hong Simon, Optican Lance M
Laboratory of Sensorimotor Research, National Eye Institute, National Institutes of Health, Bethesda, Maryland, United States of America.
PLoS One. 2008 Jul 23;3(7):e2770. doi: 10.1371/journal.pone.0002770.
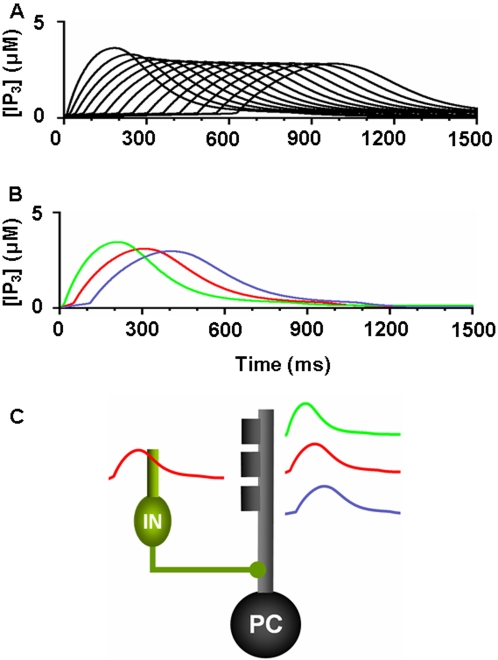
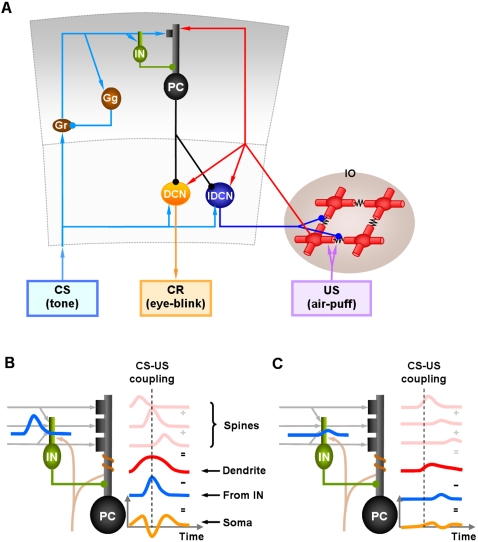
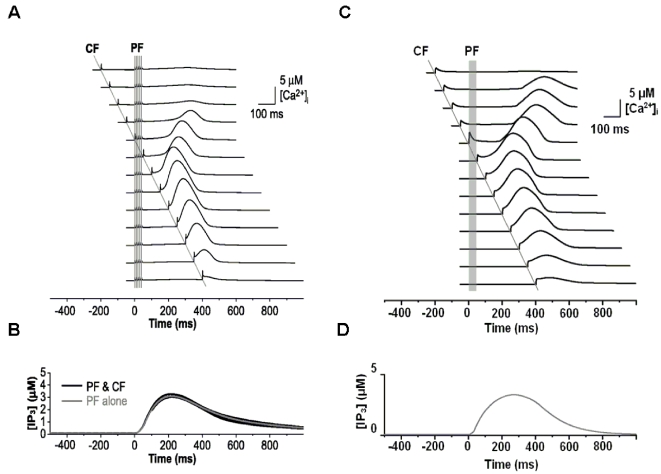
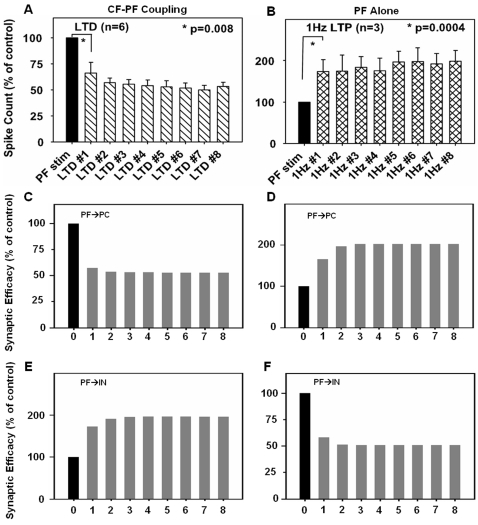
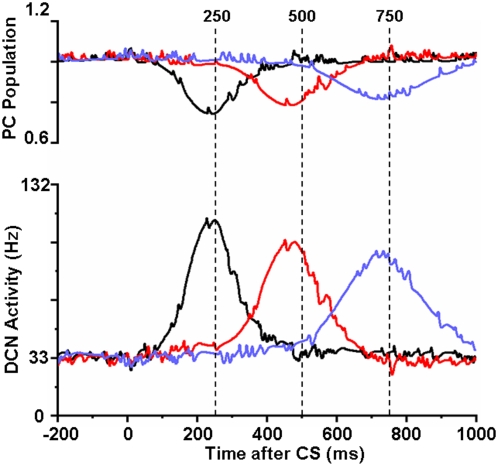
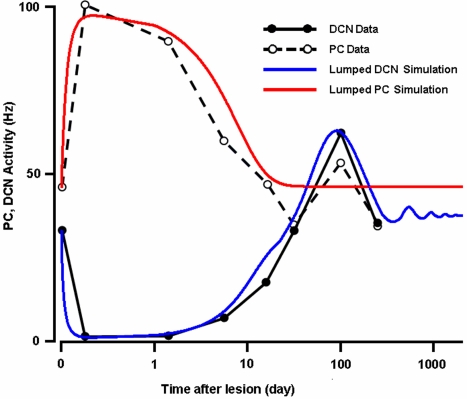
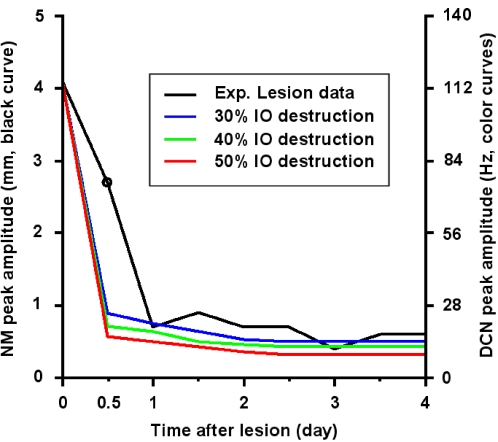
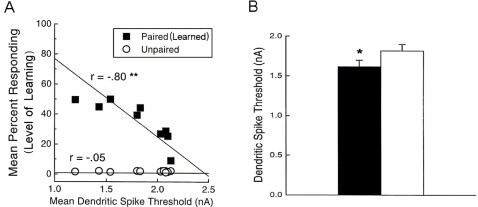
We develop a new model that explains how the cerebellum may generate the timing in classical delay eyeblink conditioning. Recent studies show that both Purkinje cells (PCs) and inhibitory interneurons (INs) have parallel signal processing streams with two time scales: an AMPA receptor-mediated fast process and a metabotropic glutamate receptor (mGluR)-mediated slow process. Moreover, one consistent finding is an increased excitability of PC dendrites (in Larsell's lobule HVI) in animals when they acquire the classical delay eyeblink conditioning naturally, in contrast to in vitro studies, where learning involves long-term depression (LTD). Our model proposes that the delayed response comes from the slow dynamics of mGluR-mediated IP3 activation, and the ensuing calcium concentration change, and not from LTP/LTD. The conditioned stimulus (tone), arriving on the parallel fibers, triggers this slow activation in INs and PC spines. These excitatory (from PC spines) and inhibitory (from INs) signals then interact at the PC dendrites to generate variable waveforms of PC activation. When the unconditioned stimulus (puff), arriving on the climbing fibers, is coupled frequently with this slow activation the waveform is amplified (due to an increased excitability) and leads to a timed pause in the PC population. The disinhibition of deep cerebellar nuclei by this timed pause causes the delayed conditioned response. This suggested PC-IN interaction emphasizes a richer role of the INs in learning and also conforms to the recent evidence that mGluR in the cerebellar cortex may participate in slow motor execution. We show that the suggested mechanism can endow the cerebellar cortex with the versatility to learn almost any temporal pattern, in addition to those that arise in classical conditioning.
我们开发了一种新模型,该模型解释了小脑如何在经典延迟眨眼条件反射中产生时间节律。最近的研究表明,浦肯野细胞(PCs)和抑制性中间神经元(INs)都具有两个时间尺度的并行信号处理流:一个是由AMPA受体介导的快速过程,另一个是由代谢型谷氨酸受体(mGluR)介导的缓慢过程。此外,一个一致的发现是,与体外研究(其中学习涉及长时程抑制,即LTD)相反,动物在自然获得经典延迟眨眼条件反射时,其PC树突(在拉塞尔小叶HVI中)的兴奋性会增加。我们的模型提出,延迟反应来自mGluR介导的IP3激活的缓慢动力学以及随之而来的钙浓度变化,而不是来自长时程增强(LTP)/长时程抑制(LTD)。到达平行纤维的条件刺激(音调)会触发INs和PC棘中的这种缓慢激活。然后,这些兴奋性信号(来自PC棘)和抑制性信号(来自INs)在PC树突处相互作用,以产生PC激活的可变波形。当到达攀爬纤维的非条件刺激(吹气)经常与这种缓慢激活耦合时,波形会被放大(由于兴奋性增加),并导致PC群体出现定时停顿。这种定时停顿对小脑深部核团的去抑制作用会引发延迟的条件反应。这种提出的PC-IN相互作用强调了INs在学习中更丰富的作用,也符合最近的证据,即小脑皮质中的mGluR可能参与缓慢的运动执行。我们表明,所提出的机制可以赋予小脑皮质学习几乎任何时间模式的通用性,除了那些在经典条件反射中出现的模式。