Kollins K M, Hu J, Bridgman P C, Huang Y Q, Gallo G
Department of Neurobiology and Anatomy, Drexel University College of Medicine, Philadelphia, Pennsylvania 19129, USA.
Dev Neurobiol. 2009 Apr;69(5):279-98. doi: 10.1002/dneu.20704.
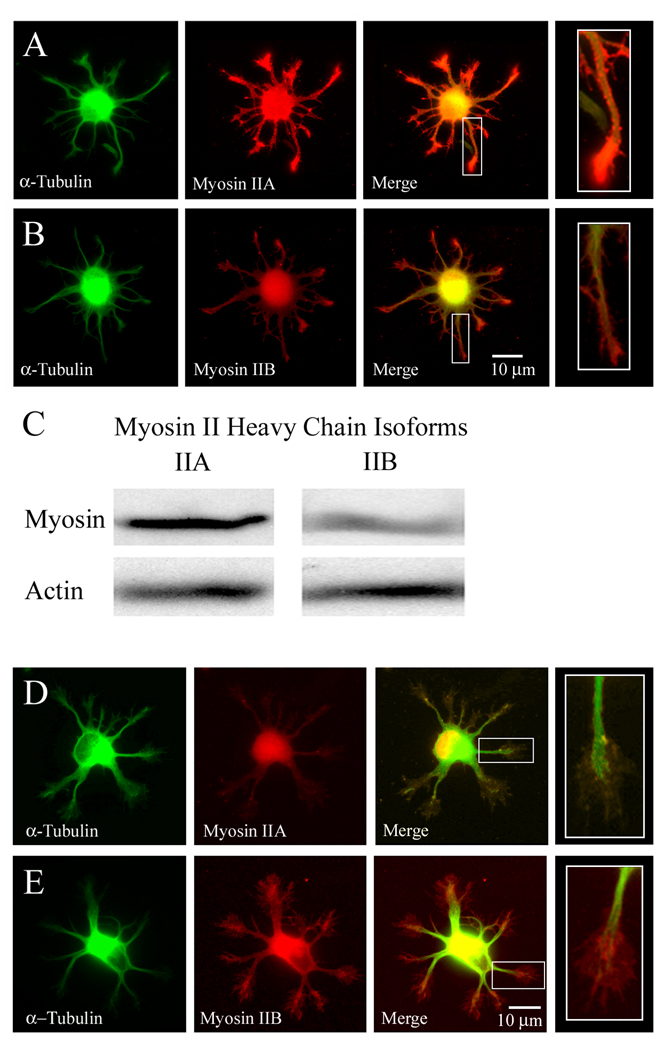
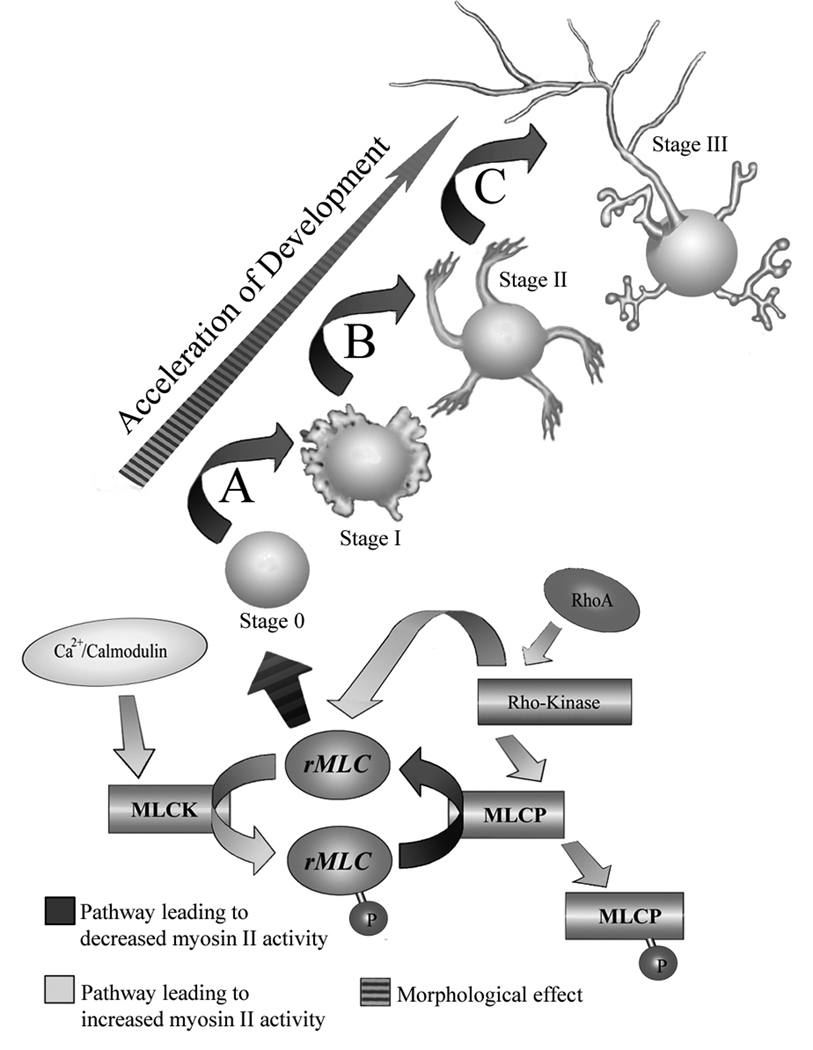
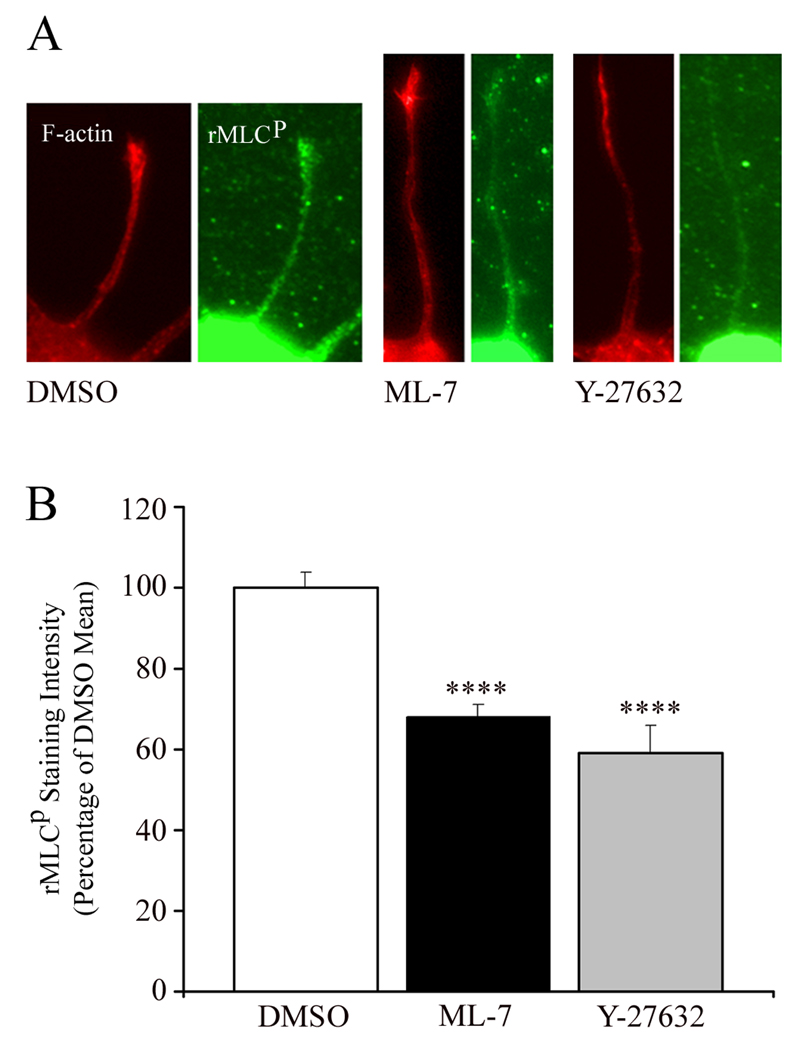
The earliest stage in the development of neuronal polarity is characterized by extension of undifferentiated "minor processes" (MPs), which subsequently differentiate into the axon and dendrites. We investigated the role of the myosin II motor protein in MP extension using forebrain and hippocampal neuron cultures. Chronic treatment of neurons with the myosin II ATPase inhibitor blebbistatin increased MP length, which was also seen in myosin IIB knockouts. Through live-cell imaging, we demonstrate that myosin II inhibition triggers rapid minor process extension to a maximum length range. Myosin II activity is determined by phosphorylation of its regulatory light chains (rMLC) and mediated by myosin light chain kinase (MLCK) or RhoA-kinase (ROCK). Pharmacological inhibition of MLCK or ROCK increased MP length moderately, with combined inhibition of these kinases resulting in an additive increase in MP length similar to the effect of direct inhibition of myosin II. Selective inhibition of RhoA signaling upstream of ROCK, with cell-permeable C3 transferase, increased both the length and number of MPs. To determine whether myosin II affected development of neuronal polarity, MP differentiation was examined in cultures treated with direct or indirect myosin II inhibitors. Significantly, inhibition of myosin II, MLCK, or ROCK accelerated the development of neuronal polarity. Increased myosin II activity, through constitutively active MLCK or RhoA, decreased both the length and number of MPs and, consequently, delayed or abolished the development of neuronal polarity. Together, these data indicate that myosin II negatively regulates MP extension, and the developmental time course for axonogenesis.
神经元极性发育的最早阶段的特征是未分化的“小突起”(MPs)的延伸,这些小突起随后分化为轴突和树突。我们使用前脑和海马神经元培养物研究了肌球蛋白II运动蛋白在MP延伸中的作用。用肌球蛋白II ATP酶抑制剂blebbistatin对神经元进行长期处理会增加MP的长度,这在肌球蛋白IIB基因敲除的情况下也能看到。通过活细胞成像,我们证明肌球蛋白II的抑制会触发小突起迅速延伸至最大长度范围。肌球蛋白II的活性由其调节轻链(rMLC)的磷酸化决定,并由肌球蛋白轻链激酶(MLCK)或RhoA激酶(ROCK)介导。对MLCK或ROCK的药理抑制会适度增加MP的长度,同时抑制这两种激酶会导致MP长度的累加增加,类似于直接抑制肌球蛋白II的效果。用细胞可渗透的C3转移酶对ROCK上游的RhoA信号进行选择性抑制,会增加MP的长度和数量。为了确定肌球蛋白II是否影响神经元极性的发育,我们在使用直接或间接肌球蛋白II抑制剂处理的培养物中检查了MP的分化。值得注意的是,抑制肌球蛋白II、MLCK或ROCK会加速神经元极性的发育。通过组成型活性MLCK或RhoA增加肌球蛋白II的活性,会减少MP的长度和数量,从而延迟或消除神经元极性的发育。总之,这些数据表明肌球蛋白II对MP的延伸和轴突发生的发育时间进程具有负调控作用。