Minko Irina G, Kozekov Ivan D, Harris Thomas M, Rizzo Carmelo J, Lloyd R Stephen, Stone Michael P
Center for Research on Occupational and Environmental Toxicology, Oregon Health & Science University, Portland, Oregon 97239, USA.
Chem Res Toxicol. 2009 May;22(5):759-78. doi: 10.1021/tx9000489.
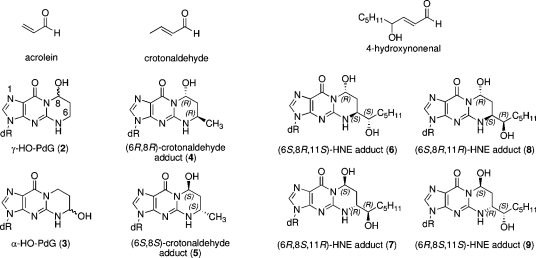
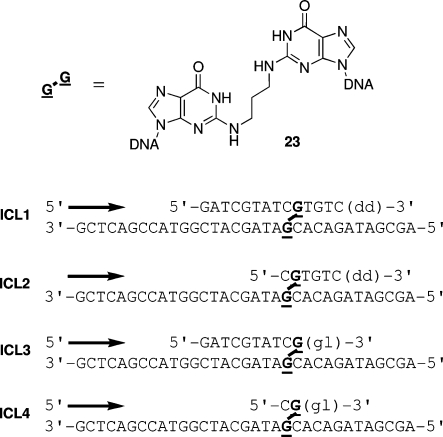
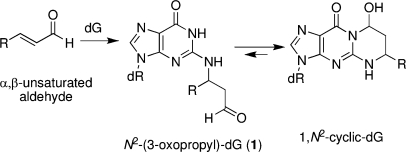
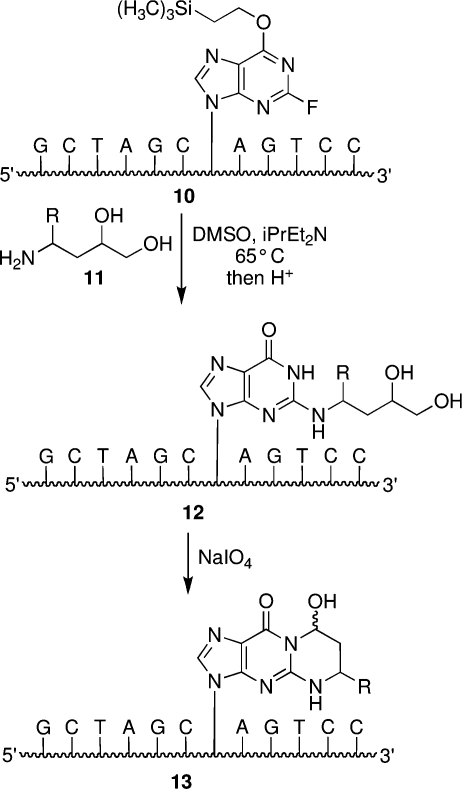
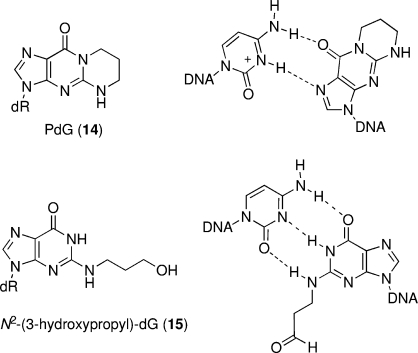
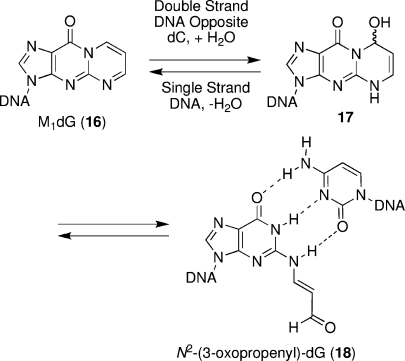
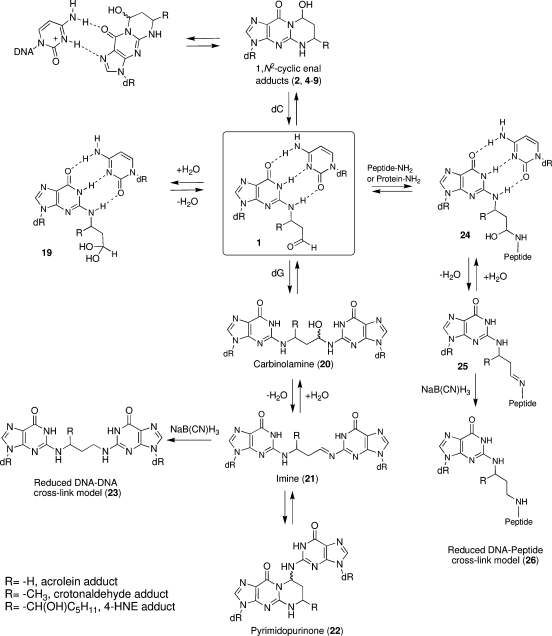
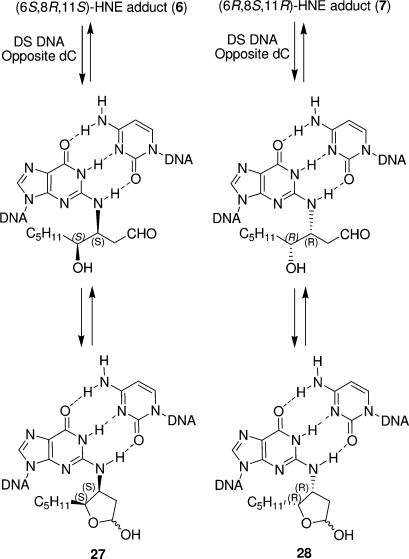
The alpha,beta-unsaturated aldehydes (enals) acrolein, crotonaldehyde, and trans-4-hydroxynonenal (4-HNE) are products of endogenous lipid peroxidation, arising as a consequence of oxidative stress. The addition of enals to dG involves Michael addition of the N(2)-amine to give N(2)-(3-oxopropyl)-dG adducts, followed by reversible cyclization of N1 with the aldehyde, yielding 1,N(2)-dG exocyclic products. The 1,N(2)-dG exocyclic adducts from acrolein, crotonaldehyde, and 4-HNE exist in human and rodent DNA. The enal-induced 1,N(2)-dG lesions are repaired by the nucleotide excision repair pathway in both Escherichia coli and mammalian cells. Oligodeoxynucleotides containing structurally defined 1,N(2)-dG adducts of acrolein, crotonaldehyde, and 4-HNE were synthesized via a postsynthetic modification strategy. Site-specific mutagenesis of enal adducts has been carried out in E. coli and various mammalian cells. In all cases, the predominant mutations observed are G-->T transversions, but these adducts are not strongly miscoding. When placed into duplex DNA opposite dC, the 1,N(2)-dG exocyclic lesions undergo ring opening to the corresponding N(2)-(3-oxopropyl)-dG derivatives. Significantly, this places a reactive aldehyde in the minor groove of DNA, and the adducted base possesses a modestly perturbed Watson-Crick face. Replication bypass studies in vitro indicate that DNA synthesis past the ring-opened lesions can be catalyzed by pol eta, pol iota, and pol kappa. It also can be accomplished by a combination of Rev1 and pol zeta acting sequentially. However, efficient nucleotide insertion opposite the 1,N(2)-dG ring-closed adducts can be carried out only by pol iota and Rev1, two DNA polymerases that do not rely on the Watson-Crick pairing to recognize the template base. The N(2)-(3-oxopropyl)-dG adducts can undergo further chemistry, forming interstrand DNA cross-links in the 5'-CpG-3' sequence, intrastrand DNA cross-links, or DNA-protein conjugates. NMR and mass spectrometric analyses indicate that the DNA interstand cross-links contain a mixture of carbinolamine and Schiff base, with the carbinolamine forms of the linkages predominating in duplex DNA. The reduced derivatives of the enal-mediated N(2)-dG:N(2)-dG interstrand cross-links can be processed in mammalian cells by a mechanism not requiring homologous recombination. Mutations are rarely generated during processing of these cross-links. In contrast, the reduced acrolein-mediated N(2)-dG peptide conjugates can be more mutagenic than the corresponding monoadduct. DNA polymerases of the DinB family, pol IV in E. coli and pol kappa in human, are implicated in error-free bypass of model acrolein-mediated N(2)-dG secondary adducts, the interstrand cross-links, and the peptide conjugates.
α,β-不饱和醛(烯醛)丙烯醛、巴豆醛和反式-4-羟基壬烯醛(4-HNE)是内源性脂质过氧化的产物,由氧化应激产生。烯醛与dG的加成涉及N(2)-胺的迈克尔加成,生成N(2)-(3-氧代丙基)-dG加合物,随后N1与醛发生可逆环化,产生1,N(2)-dG外环产物。来自丙烯醛、巴豆醛和4-HNE的1,N(2)-dG外环加合物存在于人类和啮齿动物的DNA中。烯醛诱导的1,N(2)-dG损伤在大肠杆菌和哺乳动物细胞中均由核苷酸切除修复途径修复。通过合成后修饰策略合成了含有结构明确的丙烯醛、巴豆醛和4-HNE的1,N(2)-dG加合物的寡脱氧核苷酸。已在大肠杆菌和各种哺乳动物细胞中对烯醛加合物进行了位点特异性诱变。在所有情况下,观察到的主要突变是G→T颠换,但这些加合物的错义编码能力不强。当置于与dC相对的双链DNA中时,1,N(2)-dG外环损伤会开环形成相应的N(2)-(3-氧代丙基)-dG衍生物。值得注意的是,这会在DNA的小沟中产生一个反应性醛,且加合碱基的沃森-克里克面受到适度扰动。体外复制绕过研究表明,环开损伤后的DNA合成可由聚合酶η、聚合酶ι和聚合酶κ催化。它也可以通过Rev1和聚合酶ζ依次作用的组合来完成。然而,只有聚合酶ι和Rev1这两种不依赖沃森-克里克配对来识别模板碱基的DNA聚合酶才能有效地在1,N(2)-dG闭环加合物相对位置插入核苷酸。N(2)-(3-氧代丙基)-dG加合物可发生进一步化学反应,在5'-CpG-3'序列中形成链间DNA交联、链内DNA交联或DNA-蛋白质缀合物。核磁共振和质谱分析表明,DNA链间交联包含甲醇胺和席夫碱的混合物,在双链DNA中,甲醇胺形式的连接占主导。烯醛介导的N(2)-dG:N(2)-dG链间交联的还原衍生物在哺乳动物细胞中可通过一种不需要同源重组的机制进行处理。在这些交联处理过程中很少产生突变。相比之下,还原的丙烯醛介导的N(2)-dG肽缀合物可能比相应的单加合物更具诱变性。DinB家族的DNA聚合酶,大肠杆菌中的聚合酶IV和人类中的聚合酶κ,与模型丙烯醛介导的N(2)-dG二级加合物、链间交联和肽缀合物的无错绕过有关。