Department of Microbiology and Immunology, University of Rochester Medical Center, Rochester, NY 14642, USA.
Can J Microbiol. 2009 Oct;55(10):1133-44. doi: 10.1139/w09-065.
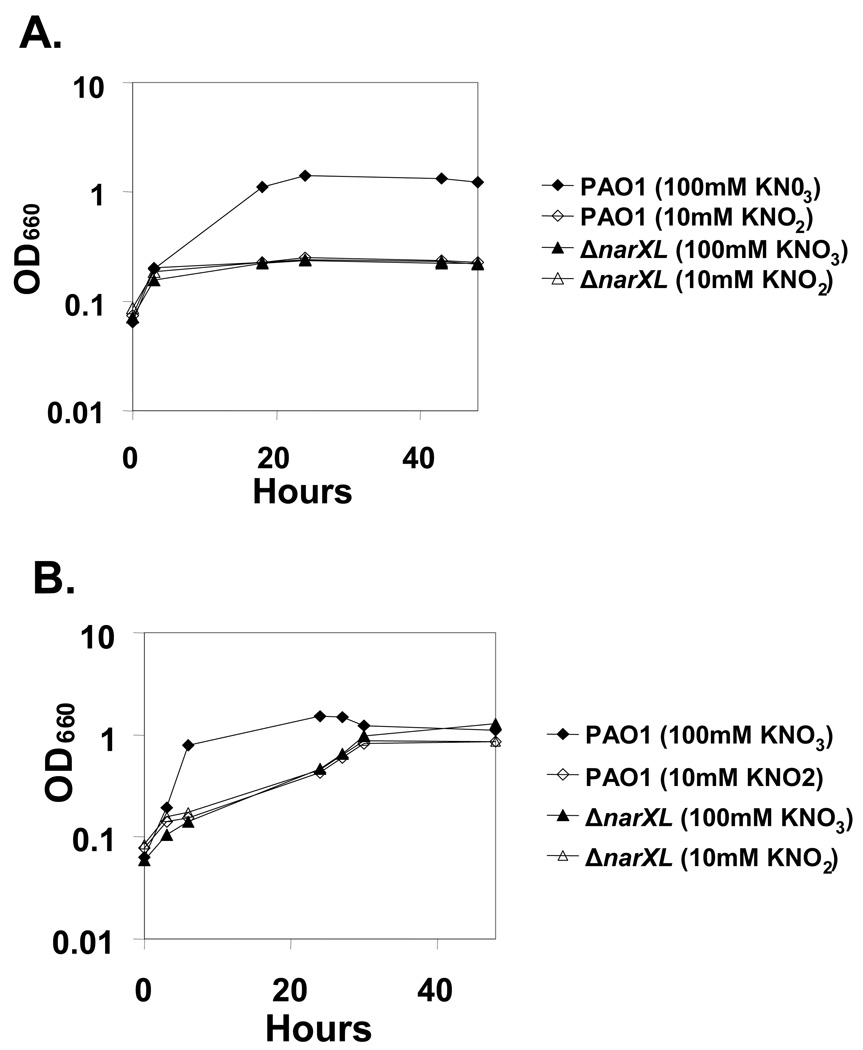
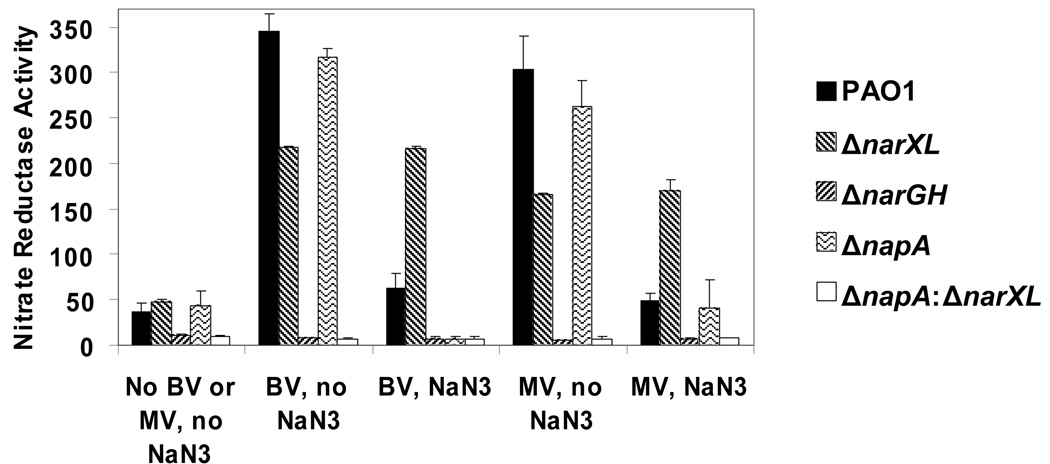
Nitrate serves as a terminal electron acceptor under anaerobic conditions in Pseudomonas aeruginosa. Reduction of nitrate to nitrite generates a transmembrane proton motive force allowing ATP synthesis and anaerobic growth. The inner membrane-bound nitrate reductase NarGHI is encoded within the narK1K2GHJI operon, and the periplasmic nitrate reductase NapAB is encoded within the napEFDABC operon. The roles of the 2 dissimilatory nitrate reductases in anaerobic growth, and the regulation of their expressions, were examined by use of a set of deletion mutants in P. aeruginosa PAO1. NarGHI mutants were unable to grow anaerobically, but plate cultures remained viable up to 120 h. In contrast, the nitrate sensor-response regulator mutant DeltanarXL displayed growth arrest initially, but resumed growth after 72 h and reached the early stationary phase in liquid culture after 120 h. Genetic, transcriptional, and biochemical studies demonstrated that anaerobic growth recovery by the NarXL mutant was the result of NapAB periplasmic nitrate reductase expression. A novel transcriptional start site for napEFDABC expression was identified in the NarXL mutant grown anaerobically. Furthermore, mutagenesis of a consensus NarL-binding site monomer upstream of the novel transcriptional start site restored anaerobic growth recovery in the NarXL mutant. The data suggest that during anaerobic growth of wild-type P. aeruginosa PAO1, the nitrate response regulator NarL directly represses expression of periplasmic nitrate reductase, while inducing maximal expression of membrane nitrate reductase.
硝酸盐在铜绿假单胞菌的厌氧条件下作为末端电子受体。硝酸盐还原为亚硝酸盐会产生跨膜质子动力,从而允许 ATP 合成和厌氧生长。内膜结合的硝酸盐还原酶 NarGHI 编码在 narK1K2GHJI 操纵子中,而周质硝酸盐还原酶 NapAB 编码在 napEFDABC 操纵子中。通过使用一组铜绿假单胞菌 PAO1 的缺失突变体,研究了这 2 种异化硝酸盐还原酶在厌氧生长中的作用及其表达的调节。NarGHI 突变体不能进行厌氧生长,但平板培养物在高达 120 h 时仍保持存活。相比之下,硝酸盐传感器-反应调节剂突变体 DeltanarXL 最初显示出生长停滞,但在 72 h 后恢复生长,并在 120 h 后达到液体培养的早期静止期。遗传、转录和生化研究表明,NarXL 突变体的厌氧生长恢复是由于 NapAB 周质硝酸盐还原酶的表达。在厌氧生长的 NarXL 突变体中鉴定到 napEFDABC 表达的新转录起始位点。此外,在新转录起始位点上游的共识 NarL 结合位点单体上进行诱变,恢复了 NarXL 突变体的厌氧生长恢复。数据表明,在野生型铜绿假单胞菌 PAO1 的厌氧生长过程中,硝酸盐应答调节因子 NarL 直接抑制周质硝酸盐还原酶的表达,同时诱导膜硝酸盐还原酶的最大表达。